 2016, Vol. 56
2016, Vol. 56
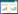


Epstein-Barr病毒的免疫调控与逃逸机制

http://dx.doi.org/10.13343/j.cnki.wsxb.20150193
中国科学院微生物研究所,中国微生物学会,中国菌物学会
中国科学院微生物研究所,中国微生物学会,中国菌物学会
文章信息
- 肖楠阳, 陈骐, 蔡少丽
- Nanyang Xiao, Qi Chen, Shaoli Cai
- Epstein-Barr病毒的免疫调控与逃逸机制
- Regulation and evasion of host immune responses by EpsteinBarr virus
- 微生物学报, 2016,56(1): 19-25
- Acta Microbiologica Sinica, 2016,56(1): 19-25
-
文章历史
- 收稿日期:2015-04-26
- 修回日期:2015-07-13
引用本文 |
肖楠阳, 陈骐, 蔡少丽. Epstein-Barr病毒的免疫调控与逃逸机制[J]. 微生物学报,2016,56(1): 19-25.
Nanyang Xiao, Qi Chen, Shaoli Cai. Regulation and evasion of host immune responses by EpsteinBarr virus[J]. Acta Microbiologica Sinica, 2016,56(1): 19-25.
Epstein-Barr病毒的免疫调控与逃逸机制
福建师范大学南方生物医学研究中心, 福建省天然免疫生物学重点实验室, 福建福州 350117
摘要: EB病毒(Epstein-Barr Virus, EBV)属于γ疱疹病毒科,是第一个被发现与人类肿瘤相关的DNA病毒。EB病毒通过激活Toll样受体(Toll like receptors, TLRs)信号通路,诱导I型干扰素的大量释放和功能性的自噬机制,从而引起机体的免疫应答。然而,相对于其他疱疹病毒, EB病毒已进化出更为精细且错综复杂的机制来破坏和逃逸宿主的免疫系统,如限制自身蛋白表达、活化宿主的泛素-蛋白酶体系统、干扰或逆转自噬与泛素化修饰等。这些机制会引发EB病毒在宿主体内的持续性感染,导致宿主免疫功能失调,引发EB病毒相关疾病(如鼻咽癌、传染性单核细胞增多症等)。因此,研究EB病毒特异性的免疫调控机制不仅对深入理解EB病毒的潜伏性感染和致癌性至关重要,而且还将为EB病毒诱发的相关疾病的免疫预防与治疗鉴定出新的潜在靶点。此文主要阐述了EB病毒调控宿主免疫应答和逃逸先天免疫应答的分子机制。
关键词:
Epstein-Barr病毒 肿瘤 TLRs 免疫逃逸
Regulation and evasion of host immune responses by EpsteinBarr virus
The Key Laboratories of Innate Inimune Biology of Fujian Province, Southern Biomedical Research Center, Fujian Normal University, Fuzhou 350117, Fujian Province, China
Abstract:Epstein-Barr virus(EBV) is the first identified human oncogenic DNA virus in the gamma-herpesvirus family. EBV triggers a cascade events of innate immune responses through Toll-like receptor signaling including the production of type I interferons and the activation of functional autophagy. However, EBV has developed much more elaborate and sophisticated strategies for subverting and escaping the host immune system, such as limiting its own gene expression, activing the host ubiquitin-specific protease system, and interfering ubiquitin modification. EBV impairs the host immune system, leading to lifelong persistent infections, which in turn result in the occurrence of EBV-associated diseases, such as nasopharyngeal carcinoma and infectious mononucleosis. Thus, to better understand the mechanisms regarding the infection latentency and oncogenicity of EBV invasion will be crucial for identifying potential immunotherapeutic targets for EBVrelated diseases, such as infectious mononucleosis and nasopharyngeal carcinoma. In this article, we discuss the research advances regarding the virology and immunology of EBV in the modulation of the host immune response and evasion.
Key words:
Epstein-Barr virus neoplasm TLRs immune evasion
Epstein-Barr病毒(EBV),又称人类疱疹病毒4型(Human herpesvirus 4,HHV-4),是第一个发现与人类肿瘤相关的DNA病毒。EB病毒在健康人群中感染普遍,约90%成人血清中可检测到EB病毒抗体[1]。作为致癌的人类肿瘤病毒之一,EB病毒对宿主免疫系统的调控与人类多种肿瘤的发生发展、特别是早期癌变密切相关,包括胃癌、肺癌、乳腺癌、鼻咽癌、霍奇金氏淋巴瘤和儿童淋巴瘤等。本实验室建立了鼻咽癌患者血浆中EB病毒DNA的检测方法,并通过real-time PCR技术检测了53例鼻咽癌患者血浆EB病毒DNA的载量,结果显示鼻咽癌患者血浆中游离的EB病毒DNA拷贝数显著高于健康人(未发表数据,相关文章正在整理中)。EB病毒主要通过唾液接触方式传播,穿过粘膜上皮屏障,感染下层淋巴组织(如扁桃体)的B细胞。在健康宿主体内,入侵的EB病毒在宿主免疫系统的监控下进入“潜伏期”,此时,病毒藏匿于淋巴细胞的细胞核内,而不表现出任何症状。在免疫功能低下的宿主体内,EB病毒于淋巴组织中大量增殖,并异位感染T细胞、NK细胞等,导致宿主免疫系统紊乱[2]。近年来,EB病毒破坏机体免疫功能的分子机制已有较大进展,但其确切的病理学和免疫学机制尚未有详尽地阐述。
1 EB病毒生物学特性EB病毒与其他的疱疹病毒相似,成熟病毒颗粒呈球形,直径约为120–180 nm,由核样物、衣壳、囊膜三部分组成。核样物主要含全长172 kb的双股线性DNA,为直径45 nm的致密物。衣壳是由162个壳微粒组成的20面体立体对称结构。囊膜由宿主细胞的核膜组成,其上含有病毒编码的膜糖蛋白,具有识别淋巴细胞上的EB病毒受体及细胞融合等功能。
EB病毒通过其GP350蛋白与B细胞CD21受体结合从而附着于B细胞。之后,EB病毒GP42蛋白与B细胞表面HLA II类分子相互作用,实现与宿主细胞膜融合。EB病毒通过细胞内吞作用进入囊泡,并在细胞质中释放出病毒颗粒。一旦病毒的蛋白质外壳被溶解,其基因组将被输送到细胞核内,并在核内利用DNA聚合酶进行自我复制[3]。
2 EB病毒侵染和活动周期EB病毒主要有潜伏感染和裂解性感染两种感染方式。EB病毒感染初始B细胞,在EB病毒核抗原2 (EBNA2)的调控下,驱动B细胞的大量增殖。通过关闭EBNA2的表达,EBV感染的细胞增殖模式转化为静息模式。EB病毒潜伏感染的静息记忆细胞离开毛囊通过输出淋巴管进入外周循环。
B细胞受EB病毒感染后,被诱导激活增殖,并触发分化,成为记忆B细胞。小部分记忆B细胞偶然性地分化为浆细胞,浆细胞经病毒裂解感染,细胞膜溶解释放出病毒颗粒,溶解的细胞会被细胞毒性T淋巴细胞(cytotoxic lymphocyte,CTL)破坏,释放的EB病毒进入外周循环,当B细胞再循环于口腔和外周组织间,处于静息状态的B细胞可能会被释放出的EB病毒感染并发生转化,导致病毒再次激活与增殖[4](图1)。

|
| 图1 EBV侵染与宿主免疫监控 Figure1 Immune monitor in EBV infection. EBV is spread by saliva contact, and then crosses epithelium to infect B cells. EBV GP350 mediates EBV entry into B cells through interaction with its B-cell receptor CD21. The virus can enhance proliferation and division of infected B cells, but this process is continuously surveilled by cytotoxic T lymphocytes (CTLs) and natural killer cell (NK). And then virus establish lifelong persistent infections in B cells in normal immunocompetent healthy hosts. These long-lived B cells are quiescent and express fewer viral protein (only EBNA1). Viral genomic DNA replicate and synthesize of specific viral proteins for packing the newly replicated DNA into infectious virions since reactivation from latency in immunocompromised hosts. |
先天免疫系统是抵御病毒感染的第一道防线,EB病毒主要通过激活Toll样受体(Toll like receptors,TLRs),诱发IFN介导的先天免疫应答。已有研究表明EB病毒原发性感染B淋巴细胞,会诱导TLR7的表达,并激活TLR7信号通路,进而介导下游信号分子IRF5的表达,促进细胞增殖。由于EB病毒基因组富含GC序列,因而成为TLR9的配体。EB病毒的感染激活了浆树突状细胞TLR9信号通路,诱导产生IFNα,并促进NK细胞和CD3+T细胞的活化[5]。
3.1.1 EB病毒GP350表面膜蛋白:大量的研究表明:EB病毒GP350这种主要的包膜糖蛋白通过与B细胞受体CD21(主要的EB病毒受体)作用进入B细胞,促使naive B 细胞转化,并大量增殖。上皮细胞和单核细胞表面的TLR2识别完整的EB病毒颗粒,而这一过程由GP350介导,表明EB病毒表面膜蛋白GP350可能作为TLR2的配体。
3.1.2 EB病毒 dUTPase结构蛋白:除了包膜蛋白GP350,由BLLF3编码的非结构蛋白dUTPase是EB病毒感染早期抗原之一,也是TLR2识别的配体[6]。TLR2识别EB病毒dUTPase蛋白,并通过依赖MyD88的信号通路激活NF-κB,诱发巨噬细胞促炎症因子TNF-α,IL-1β,IL-6,IL-10的表达。
3.1.3 EB病毒EBERs:在潜伏感染时期,EB病毒编码两个非编码和非多聚腺苷尿苷酸的小核RNA,即EBER1和EBER2。EBERs这些非编码小RNA通过分子间的碱基互补配对形成茎环状的类dsRNA结构,并由RNA聚合酶III转录表达,因此含有5’-ppp基团[7]。EBERs这些结构使EBERs成为细胞内RNA受体——维甲酸诱导基因I(RIG-I)的配体,RIG-I 识别EBERs,激活干扰素调节因子3(IRF3)并诱导IFNs,ISGs和白细胞介素IL-10的产生[8]。EBERs特别是EBER1也可通过分泌出的细胞内EBER结合蛋白La (系统性红斑狼疮相关抗原) 从EB病毒感染的细胞释放出来,由TLR3识别并激活免疫系统,诱导I型干扰素和炎症因子的产生[9]。EBERs 也结合于IFN诱导的dsRNA依赖的蛋白激酶(PKR),并抑制PKR的磷酸化作用,从而阻断PKR介导的细胞凋亡[10]。因此,EBERs可能作为多种免疫性疾病如单核细胞增多症、噬血细胞综合征的治疗靶点。
3.2 EB病毒诱导的细胞因子EB病毒早期感染过程中诱发大量I型干扰素(Interferon,IFN)的产生,促进活化自然杀伤细胞,并通过多种方式导致适应性免疫应答。研究证明EB病毒活化的T细胞和自然杀伤细胞(Natural killer cell,NK)可导致大量IFN-γ及IFN-α的产生[11]。目前,普遍认为IFN对EB病毒的感染和再激活有重要作用。EB病毒感染诱导产生的细胞炎症因子还有肿瘤坏死因子α (Tumor necrosis factor-α,TNF-α)和多种白细胞介素(Interleukin,IL)如IL-1、IL-6、IL-10、IL-2、IL-12,及免疫抑制细胞因子IL-10和转化生长因子β(Transforming growth factor-β,TGF-β)等。如在EB病毒引起的单核细胞增多症患者血清中发现有多种炎性细胞因子的增多[12]。本实验室通过IgG诱导激活EB病毒潜伏感染的Akata细胞,western检测发现细胞中核转录因子NF-κB被活化,而参与免疫反应的早期和炎症反应各阶段的许多分子包括:TNF-α、IL-1β、IL-2、IL-6、IL-8、IL-12都受NF-κB的调控,表明EB病毒可能通过TLRs激活下游NF-κB通路。
3.3 EB病毒与免疫细胞NK细胞在先天免疫系统抵御多种病原微生物包括慢性病毒感染的调控中起关键作用。NK细胞可通过释放IFN-γ抑制受EB病毒诱导转化的静息B细胞。EB病毒相关的传染性单核细胞增多症中,NK细胞数量剧增[13],且NK细胞的数目与疾病严重程度呈负相关,表明NK细胞在限制EB病毒复制的过程中发挥重要作用。在机体免疫功能正常时,在NK细胞以及CTL的监控下,病毒很少进入增殖期,但在机体免疫力下降及某些诱因的作用下,潜伏感染的EB病毒基因组可被激活而增殖导致感染。
4 EB病毒逃逸宿主免疫防御的机制大多数的病毒已进化出逃逸宿主先天免疫系统的策略,使自身在宿主细胞中成功复制增殖。95%以上的成年人都有长期且持久性的EB病毒感染,这表明EB病毒在宿主中成功规避了其免疫系统的监控。EB病毒逃逸宿主免疫系统监控的策略包括在潜伏期只表达有限的蛋白如EBNA1、EBNA3C [14]和不被T细胞监测到的非翻译编码小RNAs(EBERs和miRNAs),通过活化宿主的泛素-蛋白酶体系统、干扰或逆转自噬与泛素化修饰从而降解宿主体内抵御病毒感染的关键性蛋白。而记忆B细胞作为EB病毒长期侵染的靶细胞,并不受免疫系统监控[15]。
4.1 EB病毒核抗原EBNA1EBNA1是EB病毒在潜伏期和裂解期都能表达的唯一蛋白,能使EB病毒基因组整合到细胞内的染色体上,以使EB病毒在细胞分裂期间得以复制[16],对EB病毒在宿主细胞中的维持、潜伏感染至关重要。EBNA1含有由甘氨酸-丙氨酸编码的重复序列(即GA结构域),GA结构域可通过干扰识别阻断19S亚基的折叠功能,抑制泛素-蛋白酶的活性,避免蛋白酶的降解,阻断生成呈递I型MHC抗原的多肽,从而干扰了免疫细胞对靶细胞的识别和杀伤作用[17]。再者,EBNA1自身合成维持在较低水平,可避免CTL的识别。最近研究表明EBNA1(也包括EBNA3C和LMP1)可导致基因组的不稳定性和活性氧介导的DNA损伤[18],并促使c-myc易位,因此是潜在的致癌基因。
4.2 致瘤蛋白LMP1的研究潜伏感染膜蛋白1(LMP1)被认为是EB病毒编码的致瘤蛋白,涉及细胞转化、凋亡、分化、细胞周期以及参与恶性肿瘤的侵袭和转移[19]。EB病毒感染的细胞会分泌LMP1高表达的胞外体,并与T淋巴细胞融合,LMP1第一个跨膜区的LALLFWL肽段可直接参与并抑制T淋巴细胞的增殖和NK细胞的杀伤作用。LMP1 N端跨膜结构域与酪氨酸激酶2 (TYK2)相互作用,抑制STAT1 和STAT2的磷酸化,阻断IFN介导的抗病毒反应[20]。另一方面,LMP1诱导大量免疫调控相关基因的表达,如趋化因子CCL17 (Chemokine ligand 17)和CCL22、IL-10、IL-8和IL-6,抑制T细胞增殖,干扰分裂素诱导的IFN-γ的分泌,从而规避CTL介导的细胞免疫。此外,LMP1还通过自身高表达引起细胞自我吞噬[21],LMP1有多重的免疫调控功能,以平衡与宿主间复杂的相互作用。综上所述,EBNA1、EBNA3C、LMP1在EB病毒逃逸宿主免疫系统的分子机制中起着至关重要的作用。本实验室已建立了CRISPR/Cas9基因编辑系统,靶向受感染细胞中EB病毒核抗原EBNA1、EBNA3C及潜伏膜蛋白LMP1,定点编辑EB病毒基因组,构建重组病毒,并研究其在调控宿主免疫系统、肿瘤发生、迁移中的作用。
4.3 EB病毒miRNAs及其功能EB病毒至少含有25种由19–24 bp寡聚核苷酸组成的MicroRNAs (miRNAs),这些miRNAs靶向病毒和受感染细胞内基因,调控病毒基因的表达,以规避宿主免疫反应并诱发癌变。除了BART2 miRNA,其他miRNAs以基因簇的形式位于BART (BamHI A region rightward transcript)和BHRF (Bam HI fragment H rightward open reading frame)两个转录区域内,对这些miRNAs的功能研究表明:BART22 miRNA特异性地靶向EB病毒潜伏膜蛋白LMP2A (EB病毒表达的强免疫原性抗原),并通过下调LMP2A的表达,躲避细胞毒性T细胞的识别,以使受EB病毒感染的细胞逃逸宿主免疫系统的监控[22];BART5 miRNA可以抑制PUMA (p53-up-regulated modulator of apoptosis)(一种Bcl2家族的提前凋亡因子)的作用,从而抑制细胞凋亡,提高宿主细胞的存活率;BART2 miRNA靶向EB病毒 DNA聚合酶BALF5,并有效抑制EB病毒裂解性增殖[23]。另一方面,细胞内多种趋化因子可能作为EB病毒miRNAs靶点,从而抑制机体的免疫应答。如BHRF1-3 miRNA靶向抑制受感染的细胞表达CXCL11,从而使这部分细胞不受IFN诱导活化的T细胞攻击[24]。此外,EB病毒miRNAs还与受感染细胞的转移、分化、信号转导和抗细胞凋亡有关。miRNAs在EB病毒的发病和致癌机理中起重要作用,但大多功能未知,因而仍有待进一步研究。本实验室利用CRISPR/Cas9技术靶向编码miRNAs的BART基因簇,并构建荧光素酶表达系统,研究由BART转录的miRNAs在调控宿主免疫系统与介导EB病毒免疫逃逸中的作用。
5 问题和展望EB病毒相对其他疱疹病毒,进化出更复杂而精细的策略来调控宿主免疫系统。目前认为免疫系统中存在着一套精细的调控机制:EB病毒可严格控制潜伏期的基因表达,限制其复制增殖,以使其在免疫功能正常的宿主内始终维持在无危害状态,当免疫系统无法控制EB病毒侵染将会引发许多严重的后果。
EB病毒调控宿主免疫系统以及逃逸免疫识别是一个错综复杂的过程,更是EB病毒侵染与致瘤的关键所在。EBNA1、LMP1以及EB病毒编码的miRNA在这一多元化的过程中起着至关重要的作用,是否可以利用新型基因编辑技术——CRISPR Cas9系统用于治疗EB病毒感染引发的疾病(目前已有报道可利用CRISPR技术清除受感染细胞中整合到宿主染色体上的EB病毒基因组),以及如何以EBNA1、LMP1作为免疫治疗靶点,开发出针对这些EB病毒抗原的新型疫苗和抗病毒药物仍有待进一步研究。另外,EB病毒非编码RNAs和miRNAs在调控先天免疫中的分子机制尚未清楚,寻找宿主先天免疫系统应答EB病毒感染的关键性分子有待研究。深入理解EB病毒如何逃逸先天免疫、实现持久性的潜伏感染并诱发恶性肿瘤的机制,阐明EB病毒的发病机理,对于相应的药物开发及制定更好的免疫治疗方案以对抗包括EB病毒诱发的恶性肿瘤等疾病都具有重要的意义。
参考文献
| [1] | Su WH, Hildesheim A, Chang YS. Human leukocyte antigens and Epstein-Barr virus-associated nasopharyngeal carcinoma:old associations offer new clues into the role of immunity in infection associated cancers. Frontiers in Oncology, 2013, 3:299. |
| [2] | Sabeti M, Kermani V, Sabeti S, Simon JH. Significance of human cytomegalovirus and Epstein-Barr virus in inducing cytokine expression in periapical lesions. Journal of Endodontics, 2012, 38(1):47-50. |
| [3] | Hernádi K, Szalmás A, Mogyorósi R, Czompa L, Veress G, Csoma E, Márton I, Kónya J. Prevalence and activity of Epstein-Barr virus and human cytomegalovirus in symptomatic and asymptomatic apical periodontitis lesions. Journal of Endodontics, 2010, 36(9):1485-1489. |
| [4] | Murata T, Tsurumi T. Switching of EBV cycles between latent and lytic states. Reviews in Medical Virology, 2014, 24(3):142-153. |
| [5] | Younesi V, Nikzamir H, Yousefi M, Khoshnoodi J, Arjmand M, Rabbani H, Shokri F. Epstein Barr virus inhibits the stimulatory effect of TLR7/8 and TLR9 agonists but not CD40 ligand in human B lymphocytes. Microbiology and Immunology, 2010, 54(9):534-541. |
| [6] | Ariza ME, Glaser R, Kaumaya PTP, Jones C, Williams MV. The EBV-encoded dUTPase activates NF-κB through the TLR2 and MyD88-dependent signaling pathway. The Journal of Immunology, 2009, 182(2):851-859. |
| [7] | Ablasser A, Bauernfeind F, Hartmann G, Latz E, Fitzgerald KA, Hornung V. RIG-I-dependent sensing of poly(dA:dT) through the induction of an RNA polymerase III-transcribed RNA intermediate. Nature Immunology, 2009, 10(10):1065-1072. |
| [8] | Samanta M, Iwakiri D, Takada K. Epstein-Barr virus-encoded small RNA induces IL-10 through RIG-I-mediated IRF-3 signaling. Oncogene, 2008, 27(30):4150-4160. |
| [9] | Iwakiri D, Zhou L, Samanta M, Matsumoto M, Ebihara T, Seya T, Imai S, Fujieda M, Kawa K, Takada K. Epstein-Barr virus(EBV)-encoded small RNAs is released from EBV-infected cells and activates signaling from toll-like receptor 3. The Journal of Experimental Medicine, 2009, 206(10):2091-2099. |
| [10] | Ruf IK, Lackey KA, Warudkar S, Sample JT. Protection from interferon-induced apoptosis by Epstein-Barr virus small RNAs is not mediated by inhibition of PKR. Journal of Virology,2005, 79(23):14562-14569. |
| [11] | Quan TE, Roman RM, Rudenga BJ, Holers VM, Craft JE. Epstein-Barr virus promotes interferon-α production by plasmacytoid dendritic cells. Arthritis & Rheumatism, 2010, 62(6):1693-1701. |
| [12] | Kimura H, Ito Y, Kawabe S, Gotoh K, Takahashi Y, Kojima S, Naoe T, Esaki S, Kikuta A, Sawada A, Kawa K, Ohshima K, Nakamura S. EBV associated T/NK-cell lymphoproliferative diseases in nonimmunocompromised hosts:prospective analysis of 108 cases. Blood, 2012, 119(3):673-686. |
| [13] | Heuts F, Rottenberg ME, Salamon D, Rasul E, Adori M, Klein G, Klein E, Nagy N. T cells modulate Epstein-Barr virus latency phenotypes during infection of humanized mice. Journal of Virology, 2014, 88(6):3235-3245. |
| [14] | Juno JA, Keynan Y, Fowke KR. Invariant NKT cells:regulation and function during viral infection. PLoS Pathogens, 2012, 8(8):e1002838. |
| [15] | Allan LL, Stax AM, Zheng DJ, Chung BK, Kozak FK, Tan RS, van den Elzen P. CD1d and CD1c expression in human B cells is regulated by activation and retinoic acid receptor signaling. The Journal of Immunology, 2011, 186(9):5261-5272. |
| [16] | Fiola S, Gosselin D, Takada K, Gosselin J. TLR9 contributes to the recognition of EBV by primary monocytes and plasmacytoid dendritic cells. The Journal of Immunology, 2010, 185(6):3620-3631. |
| [17] | Unterholzner L, Bowie AG. The interplay between viruses and innate immune signaling:recent insights and therapeutic opportunities. Biochemical Pharmacology, 2008, 75(3):589-602. |
| [18] | Gruhne B, Sompallae R, Masucci MG. Three Epstein-Barr virus latency proteins independently promote genomic instability by inducing DNA damage, inhibiting DNA repair and inactivating cell cycle checkpoints. Oncogene, 2009, 28(45):3997-4008. |
| [19] | Ersing I, Bernhardt K, Gewurz BE. NF-κB and IRF7 pathway activation by Epstein-Barr virus latent membrane protein 1. Viruses, 2013, 5(6):1587-1606. |
| [20] | Geiger TR, Martin JM. The Epstein-Barr virus-encoded LMP-1 oncoprotein negatively affects Tyk2 phosphorylation and interferon signaling in human B cells. Journal of Virology, 2006, 80(23):11638-11650. |
| [21] | Pratt ZL, Zhang JZ, Sugden B. The latent membrane protein 1(LMP1) oncogene of Epstein-Barr virus can simultaneously induce and inhibit apoptosis in B cells. Journal of Virology, 2012, 86(8):4380-4393. |
| [22] | Kalla M, Hammerschmidt W. Human B cells on their route to latent infection-early but transient expression of lytic genes of Epstein-Barr virus. European Journal of Cell Biology, 2012, 91(1):65-69. |
| [23] | Shinozaki-Ushiku A, Kunita A, Isogai M, Hibiya T, Ushiku T, Takada K, Fukayama M. Profiling of virus-encoded microRNAs in Epstein-Barr virus-associated gastric carcinoma and their roles in gastric carcinogenesis. Journal of Virology, 2015, 89(10):5581-5591. |
| [24] | Kuzembayeva M, Hayes M, Sugden B. Multiple functions are mediated by the miRNAs of Epstein-Barr virus. Current Opinion in Virology, 2014, 7:61-65. |