 2021, Vol. 61
2021, Vol. 61中国科学院微生物研究所,中国微生物学会,中国菌物学会
文章信息
- 李文骥, 郑平. 2021
- Wenji Li, Ping Zheng. 2021
- 硫氧化细菌源单质硫的生成、转运和回收
- Formation, transport and recovery of biogenic elemental sulfur from sulfur-oxidizing bacteria
- 微生物学报, 61(1): 25-40
- Acta Microbiologica Sinica, 61(1): 25-40
-
文章历史
- 收稿日期:2020-03-05
- 修回日期:2020-05-14
- 网络出版日期:2020-05-19


引用本文 |



硫元素在全球生态系统中广泛存在,是生物生长所必需的营养元素。硫元素存在于-2价和+6价之间的多种含硫化合物中。常见的还原态含硫化合物有H2S、硫化物、硫代硫酸盐(S2O32-)、亚硫酸盐(SO32-)等[1]。硫氧化细菌(Sulfur-oxidizing bacteria,SOB)能通过复杂的代谢途径将这些还原态含硫化合物氧化为硫酸盐。
单质硫是还原态含硫化合物氧化过程中的中间产物,通常的赋存形式为硫粒。据研究,大多数光能营养型绿色硫细菌(Green sulfur bacteria,GSB)和部分化能营养型SOB (如Thiobacillus)可将单质硫排至胞外。反之,一些SOB (如Allochromatium vinosum,Thiomargarita namibiensis)不具有外排单质硫的能力,可在胞内积累大量硫粒[2-4]。迄今为止,还未获得一个适用于不同种类硫粒生成和氧化的普适机理。目前虽有很多关于单质硫组成、结构和胞内分布的文献,但鲜有关于单质硫跨膜转运的报道。此外,SOB菌种、培养条件、基质种类等都是单质硫生成的决定因素,值得深入研究。
经过加工,生物源单质硫可转变为高品质硫磺[5-6]。在工业上,硫磺可用于溶剂、表面活性剂、油品添加剂等产品的生产;在农业上,硫磺可用于饲料、硫肥、农药等产品的生产;在医药上,硫磺可用于硫胺、青霉素、甲硝唑等产品的生产。鉴于单质硫的应用潜力和经济价值,从生物脱硫系统中高效回收单质硫资源具有重要意义。
本文拟结合现有的文献报道和本课题组的前期研究成果,对SOB源单质硫的生成过程、赋存形式、代谢特性、转运机理及生物脱硫系统中单质硫的回收等方面进行综述,以期为泌硫型生物脱硫工艺的研创提供参考。
1 硫氧化细菌源单质硫的生成 1.1 硫氧化代谢途径硫化物氧化为硫酸盐的代谢途径可视为两步反应(S2-→S0和S0→SO42-),单质硫是中间产物[7-9]。单质硫生成涉及两种酶:Sulfide:quinone reductase (Sqr)和Flavocytochrome c sulfide dehydrogenase (Fcc)[10-12]。Sqr和Fcc酶的作用位点均分布于SOB的周质空间内,目前已鉴定的Sqr酶都是膜结合单亚基黄素蛋白[13],Fcc酶则呈可溶性周质蛋白和膜结合蛋白两种形式[14]。在硫化物氧化为单质硫的过程中,先由Fcc酶将e-传递给氧化态细胞色素c(cyt+)(式1)[15];再由细胞色素c氧化酶催化还原态细胞色素c(cyt)的氧化反应和O2的还原反应(式2),回补cyt+[16]。因为细胞色素c氧化酶对高浓度HS-敏感,所以在高浓度硫化物条件下,Fcc酶催化系统易因cyt+匮乏而被抑制[10, 17-18]。与此相反,在Sqr酶催化系统中,不以细胞色素c作为电子载体,其不受高浓度硫化物的影响[11, 19]。据推测,在SOB中,可能同时兼有上述两种酶系统,并由环境条件决定优势酶及其活性[11]。
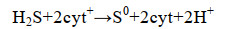
|
公式(1) |
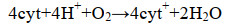
|
公式(2) |
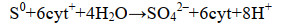
|
公式(3) |
单质硫可进一步氧化为硫酸盐,其代谢路径由多种酶促反应构成,均通过细胞色素c作为电子传递体(式3)[20]。在Dissimilatory sulfite reductase (Dsr)的作用下,单质硫氧化为亚硫酸盐;亚硫酸盐再进一步氧化为硫酸盐[20-21]。Dsr酶是硫循环中氧化环节和还原环节的关键酶,既可在厌氧呼吸条件下催化亚硫酸盐转化为硫化物,也可在硫化物氧化过程中催化单质硫逆向转化为亚硫酸盐[22]。本课题组的研究结果表明,sqr和dsrA基因的丰度和转录水平分别能表征“短程硫氧化”(S2-→S0)和“全程硫氧化”(S0→SO42-)活性,单质硫的积累可通过强化“短程硫氧化”并抑制“全程硫氧化”实现[8]。当硫化物供不应求时,化能自养型SOB能以单质硫作为能源,将其氧化为硫酸盐,释放能量用以维持生命活动[1, 23]。
1.2 单质硫的生成硫化物通过自由扩散由胞外进入SOB的周质空间[13-14, 24-25]。在硫化物氧化为单质硫的过程中,Sqr和Fcc酶起着关键作用。Sqr酶是一种膜结合周质蛋白,分布于细胞壁内膜上,酶活性中心位于周质空间内[15, 24]。Fcc酶系统由黄素蛋白亚基(FccB)和细胞色素c亚基(FccA)构成,分布于周质空间内或结合在膜上[25-26]。Sqr和Fcc均能将硫化物氧化为多硫化物,但多硫化物进一步转化为单质硫的机制仍未明晰[13-14, 27-28]。Zhang等(2014)[29]通过研究Acidithiobacillus ferrooxidans中Sqr催化的硫化物氧化反应,提出了一种单质硫生成机制(图 1)。当一个硫离子与Cys356的Sγ相互作用时,FAD获得2个电子,硫原子连接到多硫键上。A. ferrooxidans中Sqr的活性位点包括2个半胱氨酸(Cys160,Cys356),它们参与电子向FAD的转移和多硫键的形成,第3个半胱氨酸(Cys128)则与产物的释放有关[30-31]。

Steudel等(1996)[32]曾提出硫化物化学氧化为环辛硫(S8)的机理,推测SOB也可能以类似的方式对硫化物进行生物氧化。在这一机制中,HS-阴离子被氧化为硫化物自由基(HS·或S·-)(式4-5)。
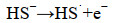
|
公式(4) |
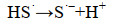
|
公式(5) |
这些自由基是系列复杂反应的基础,包括离子的自由基化和自由基离子的二聚化,产生多硫化物阴离子(Sn2-)(式6-16)。
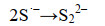
|
公式(6) |
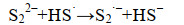
|
公式(7) |
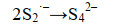
|
公式(8) |
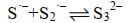
|
公式(9) |
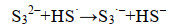
|
公式(10) |
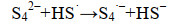
|
公式(11) |
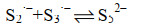
|
公式(12) |
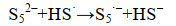
|
公式(13) |
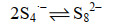
|
公式(14) |
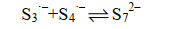
|
公式(15) |
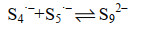
|
公式(16) |
多硫化物阴离子与氢离子反应,形成单质硫(式17-19)。
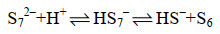
|
公式(17) |
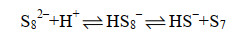
|
公式(18) |
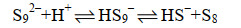
|
公式(19) |
一类SOB可将硫粒转运至胞外,即泌硫型SOB如Chlorobiaceae科的GSB、Ectothiorhodospiraceae科的紫色硫细菌(Purple sulfur bacteria,PSB)、紫色非硫细菌(Purple nonsulfur bacteria,PNSB)以及Thiobacillus等属的无色硫细菌(Colorless sulfur bacteria,CSB)[3]。另一类则可将硫粒贮存于胞内,即贮硫型SOB如Beggiatoa和Thioploca菌种[2]。
1.3.1 胞内硫粒:据文献报道,胞内硫粒的表面都有蛋白质包囊,且可利用冷冻蚀刻技术和透射电镜技术观察到[2],本课题组观察到的胞内硫粒表面具有明显的包囊结构(图 2-A)。目前,对于胞内硫粒蛋白质包囊的认知,大多来自Chromatiaceae科PSB型SOB的研究。Schmidt等(1971)[33]通过离心得到胞内硫粒,用氯仿提取,分离单质硫和蛋白质包囊,发现Allochromatium vinosum蛋白质包囊由13.5 kDa蛋白组成。Imhoff等(1998)[34]则发现,Isochromatium buderi的蛋白质包囊由18.5 kDa蛋白组成。当SOB生长在含硫化物的培养基中时,Beggiatoa胞内硫粒的外表面也有15 kDa蛋白[35]。以Allochromatium vinosum为例,进一步用高效液相色谱分析蛋白质包囊,发现13.5 kDa蛋白质由2个10.6 kDa的同源亚单元(SgpA和SgpB)组成。胞内硫粒蛋白质包囊中还有第3种8.5 kDa的亚单元(SgpC)[36]。三种蛋白质共同构成了Allochromatium vinosum胞内硫粒的蛋白质包囊(图 2-B)[2]。当编码SgpA蛋白的基因失活时,胞内硫粒仍能生成,表明两个同源蛋白(SgpA或SgpB)中只有一个是必需的[37]。与野生型相比,sgpA基因或sgpB基因的失活没有使SOB表现出任何差异,而sgpC基因的失活则会使SOB形成较小的胞内硫粒。sgpB和sgpC双基因突变体,则不能以硫化物为基质进行生长,也无法形成胞内硫粒,表明蛋白质包囊是形成胞内硫粒的必要条件[38]。

利用X射线能量色散分析技术对Beggiatoa alba的细胞内含物进行鉴定,证明内含物主要由硫元素构成,但不能确定是链状多硫化物还是环辛硫[39]。而X射线吸收近边结构(X-ray absorption near-edge structure,XANES)光谱是鉴定胞内硫粒更加有效的技术[2]。通过XANES鉴定发现,PSB以硫化物为基质进行光合自养生长时,胞内硫粒以有机多硫化物链状结构(R-Sn-R1)存在,一端或两端为有机基团,且未检测出环状硫分子[40]。而在Beggiatoa和Thioploca菌细胞中,胞内硫粒则以环辛硫为主[41]。
以Beggiatoa为例,胞内硫粒在周质空间内生成,胞内硫粒被细胞膜包裹并内陷,SOB的细胞结构和胞内硫粒的位置分布如图 3-C所示[42]。Strohl等(1981)研究了Beggiatoa alba strain B15LD胞内硫粒的结构特征(图 3-A、B),细胞膜与硫粒清晰可辨,尽管胞内硫粒已深入细胞质内部,但其仍然处于延伸的周质空间内[43-44]。

1.3.2 胞外硫粒:
Janssen等(1996)[45]通过扫描电镜观察实验室生物脱硫系统中Thiobacillus SOB细胞表面,发现了胞外硫粒(图 4-A)。Thiorhodospira sibirica (图 4-B)[3]和Thermoanaerobacter sulfurigignens (图 4-C)[46]也可将单质硫排至细胞外。在本课题组运行的BISURE(BIological SUlfide REmoval)生物脱硫系统中,通过扫描电镜结合能谱分析,同样观察到了SOB表面的胞外硫粒(图 4-D)[8]。

|
| 图 4 Thiobacillus(A)[45]、Thiorhodospira sibirica(B)[3]、Thermoanaerobacter sulfurigignens(C)[46]的胞外硫粒和本课题组SOB胞外硫粒(绿点) (D)的能谱分析 Figure 4 Extracellularly stored sulfur of Thiobacillus (A)[45], Thiorhodospira sibirica (B)[3]and Thermoanaerobacter sulfurigignens (C)[46] and EDS analysis of the particulate substance (green dots) outside the SOB in our study (D). |
与胞内硫粒相比,胞外硫粒表面具有亲水性,覆盖在表面的复合带电层由一层多硫化物和有机物构成[3]。在以连四硫酸盐为基质进行好氧生长时,Acidithiobacillus ferrooxidans可在胞外产生具有17个硫原子的链状多硫化物,并通过疏水作用附着在以环辛硫为主的内核上,具有疏水的内核和亲水的表面[3, 41, 47]。利用X射线衍射分析Thiobacillus sp.产生的胞外硫粒,发现其主要成分
为正交硫晶体[4]。化学成因正交硫晶体在水中的溶解度仅为5 μg/L[48],几乎不能溶解于水,而微生物成因单质硫可在水中溶解。胞外硫粒的亲水性可归因于其表面物质的亲水性[4]。据文献报道,疏水性的化学硫可在Thiobacillus thiooxidans作用下被水润湿,这是因为在颗粒表面形成了有机表面活性物质[49-50]。但迄今为止,未见有关胞外硫粒表面蛋白质包囊的文献报道。
2 硫氧化细菌源单质硫的转运单质硫的前体物质多硫化物形成于周质空间,而胞外硫粒的产生是一大类SOB代谢过程中的普遍现象。对于硫氧化细菌源单质硫的转运机理,至今仍无定论。根据已有文献报道,本文总结了3种单质硫转运机理,如图 5所示。

|
| 图 5 硫氧化细菌源单质硫的3种转运机理 Figure 5 Three transport mechanisms of biogenic elemental sulfur from sulfur-oxidizing bacteria. Sqr: sulfide: quinone reductase; Fcc: flavocytochrome c sulfide dehydrogenase; Corg: dissolved organics; Td: thiosulfate dehydrogenase; Tth: tetrathionate hydrolase. |
2.1 胞外间接生成
Marnocha等(2016)[51]和Hanson等(2016)[52]均利用冷冻扫描电镜发现,Chlorobaculum tepidum能在细胞外一定距离处产生胞外硫粒(图 6-A),并且在胞外硫粒生成和氧化过程中均检测到了多硫化物。基于实验结果,提出了一种Cba. tepidum生成和氧化胞外硫粒的模型,其中溶解性多硫化物被认为是关键中间产物,如图 6-B所示。

Cosmidis等(2019)[53]报道了一种用于解析单质硫形成与稳定的“S0有机矿化(S0 organomineralization)”反应,基于该反应机理,Cron等(2019)[54]利用化能自养型Sulfuricurvum kujiense进行实验,结果表明S. kujiense转运至胞外的可溶性有机物对胞外硫粒的形成和稳定是必需的,即图 5中第2种机理。一部分在周质空间产生的多硫化物扩散到胞外,生成环辛烷;另一部分多硫化物与可溶性有机物在胞外形成有机多硫化物,并在单质硫表面自组装形成有机包囊。SOB利用可溶性有机物,提高了胞外生物可利用单质硫的稳定性,但其微生物机理尚未明晰。
虽然胞内产生的多硫化物可以转运至胞外生成单质硫,但单质硫仅能在非常有限的环境条件中(-0.05V < Eh < 1.5V和pH < 5)维持热力学稳定[55]。Marnocha等(2019)[56]研究发现,Chlorobaculum tepidum产生的单质硫明显有别于化学硫,它可被由蛋白质和多糖构成的有机包囊包裹(图 6-C),使其表面具有亲水性,并减缓其向结晶态转化。大量研究表明,单质硫结晶程度越高,生物可利用性越低[52, 57]。因此,有机包囊能缓解单质硫的老化和结晶,或许可保障SOB更加有效地利用生物源单质硫。
Beard等(2011)[58]研究发现,在限制氧气供应的条件下,Acidithiobacillus ferrooxidans ATCC 23270能在硫代硫酸盐培养基上生长,并产生胞外硫粒。硫代硫酸盐在周质空间内被氧化为连四硫酸盐并扩散至胞外,在胞外酶——四硫化物水解酶(tetrathionate hydrolase, Tth)的作用下,经过一系列缩合反应(图 7),最终生成胞外硫粒,即图 5中第3种机理。检测到的胞外Tth活性为Steudel等(1987)[59]提出的由Tth驱动的亲水性硫粒生成模型提供了证据支持。该研究首次提出一种基于S4I途径(硫代硫酸盐代谢途径)的胞外硫粒生成机制。

2.2 胞内直接外泌
外膜囊泡(Outer-membrane vesicles,OMVs)是细菌细胞壁外膜包裹周质空间内容物后形成的球形囊泡。通常,只有革兰氏阴性细菌才有周质空间和细胞壁外膜[60]。在革兰氏阴性菌中,OMVs的直径约为20-250 nm,由外膜脂质、外膜蛋白质和周质空间内容物等构成[61]。OMVs有多种功能,其中之一是排出周质空间中的错叠蛋白、肽聚糖片段或异常脂多糖等。细胞壁外膜包裹周质空间内容物后形成球形囊泡,向细胞外凸出并缢断脱落,排出细胞不需要的成分[62-63]。迄今为止,未见有关周质空间内单质硫跨膜外泌过程与机理的文献报道。
基于本课题组的前期研究成果[8],有3个有力证据支持SOB的泌硫机理与OMVs有关。(1) SOB是革兰氏阴性细菌,革兰氏阴性细菌产生OMVs是普遍现象;(2) SOB在周质空间内形成单质硫作为储备能源,硫化物供应充足时,无需再储存单质硫则需分泌到胞外,而OMVs恰恰具有此类运输功能;(3)透射电镜结果显示,紧靠菌体外部可见许多球形囊泡结构,其直径均在200 nm以下(图 8-A),恰好落在OMVs的直径范围(20-250 nm)内。基于以上分析,本课题组提出了“外膜囊泡裹运”的泌硫机理假设,即图 5中第1种机理。幸运的是,本课题组在同一个SOB菌体中看到了“外膜囊泡裹运”的整个泌硫过程,结果如图 8-B所示。步骤1:单质硫积累于周质空间内,形成直径约为200 nm的硫粒,细胞壁外膜被硫粒顶起,形成向胞外凸出的囊泡,囊泡将硫粒包裹于其中;步骤2:囊泡基部的细胞壁外膜内陷缢断,囊泡携带包裹的单质硫脱离细胞;步骤3:囊泡脱离细胞后,细胞壁外膜上的缢断部位破损,通过合成作用进行外膜修补,修复的细胞壁外膜不再平整,向胞外凸出形成隆起,不妨称之为“泌硫痕”。基于以上假设,本文提出了“外膜囊泡裹运”泌硫机理的结构模型,如图 8-C所示。但是,该泌硫机理是基于本课题组BISURE反应系统中混培菌而非SOB纯培菌提出的,因此,SOB泌硫机理还有待以纯培菌深入研究。

|
| 图 8 “外膜囊泡裹运”泌硫机理假设[8] Figure 8 "OMVs-dependent" sulfur-secreting hypothesis[8]. A: spherical vesicles outside SOB (red circle); B: "OMVs-dependent" sulfur-secreting process; S-S0 (blue arrows), OM-Outer-membrane (red arrows), PS-Periplasmic space (green arrows); C: proposed structural model for "OMVs-dependent" sulfur secretion. |
3 硫氧化细菌源胞外单质硫的回收
进入生物脱硫系统的硫化物有三种命运:一是氧化为单质硫后贮存于细胞内;二是氧化为单质硫后分泌于细胞外;三是氧化为硫酸分泌于细胞外。由于现有生物脱硫系统大多采用混培菌群,再加上脱硫系统各区域的异质性,受不同O/S比(溶解氧与硫化物的摩尔比)的影响,细胞贮硫和细胞泌硫皆有发生。硫化物氧化成硫酸,会造成酸污染;硫化物氧化成单质硫并贮存于细胞内,则不利于单质硫的分离回收,尤其在沼气生物脱硫系统中,易将含单质硫细胞随吸收液携带到H2S吸收系统中,堵塞装有填料的H2S吸收塔系统[64]。因此,要实现胞外单质硫的回收利用,SOB既需有高反应能力,也需有泌硫能力。
3.1 单质硫的得率SOB氧化硫化物的化学计量式如式(11)和(12)所示。理论上,O/S比为1︰2时,硫化物全部氧化为单质硫;O/S比为2︰1时,硫化物全部氧化为硫酸盐;O/S比不同,硫化物氧化产物也不同[23]。有研究表明,通过精准供氧,可提高对生物源单质硫的选择性。研究表明,在限制供氧(O/S比为0.5-1.5)的条件下,SOB可将绝大部分硫化物氧化为单质硫;而当O/S比 > 1.5时,氧化产物以硫酸盐为主,单质硫得率较低[1]。但只要以O2作为电子受体,硫化物的化学氧化反应不可避免,必然产生硫代硫酸盐[10]。当O/S比低于0.6时,硫化物生物氧化反应受到抑制,化学氧化反应占据主导地位,氧化产物以硫代硫酸盐为主。然而,一味地降低O/S比抑制硫酸盐的产生也不是最佳选择[65-67]。研究表明,在没有硫酸盐生成的条件下,SOB生长受到抑制,无法实现自维持,因为单质硫进一步氧化为硫酸盐所释放的能量是SOB合成代谢所必需的[66-68](式11-13)。
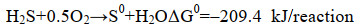
|
公式(11) |
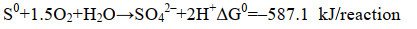
|
公式(12) |
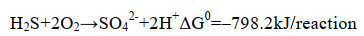
|
公式(13) |
在硫化物浓度相对稳定的连续流反应器中,溶解氧(Dissolved oxygen,DO)控制是控制硫化物氧化产物得率的有效方法之一[69-71]。据文献报道,当DO浓度控制在0.1 mg/L左右时,单质硫得率可达90 mol%以上[69]。然而,由于硫化物在反应系统中的持续消耗,系统中DO的动态性导致O/S比很难维持恒定[72-73]。作为一种替代,氧化还原电位(Oxidation-reduction potential,ORP)由硫化物浓度和溶解氧共同决定,已被证明是更可行的控制硫化物氧化反应的指标。据文献报道,在实现高效硫化物去除和单质硫回收的条件下,最优ORP在-400 mV至-137 mV之间,具有较大波动范围(表 1),这可能是由于SOB、培养基组成和生化反应的不同所致。鉴于ORP控制的普适性,它被广泛应用于沼气生物脱硫和天然气加工过程中,用以实现硫化物的脱除和单质硫的回收[23]。
| Factor | Value | Dominant final product | Yield/% | References |
| O/S ratio | 0.65 | S0 | 83.3±0.7 | [66] |
| 0.65 | S0 | 85-88 | [67] | |
| 0.63±0.04 | S0 | 92.2±0.9 | [11] | |
| 0.6-1.0 | S0 | 73±10 | [65] | |
| 0.5 | S0 | 85 | [74] | |
| DO/(mg/L) | 0.1-0.15 | S0 | — | [70] |
| 0.1 | S0 | 90 | [71] | |
| 1 | S0 | 83 | [69] | |
| ORP/mV | -100-80 | S0 | 90 | [71] |
| -147-137 | S0 | 90 | [72] | |
| -400-300 | S0 | 65 | [73] | |
| -400-360 | S0 | — | [76] | |
| -: no results. | ||||
尽管通过精准供氧可提高单质硫得率,但仍有部分硫化物被氧化为硫酸盐和硫代硫酸盐。在现有的大部分生物脱硫系统中,生物源单质硫的得率为(80-90) mol%,进一步提升单质硫得率则遇到瓶颈[66-67, 74]。在生物脱硫系统中,氧化产物主要是单质硫、硫酸盐和硫代硫酸盐三种,有效抑制硫酸盐和硫代硫酸盐的生成,成为优化单质硫得率的关键。Klok等(2019)[10]通过在沼气喷淋塔与好氧反应器之间增设厌氧反应器,使生物源单质硫得率由对照组(无厌氧反应器)的75.6 mol%大幅提升至96.6 mol%。在厌氧反应器内,高浓度硫化物抑制了以细胞色素c作为电子载体的硫酸盐生成反应,从而降低了硫酸盐得率;在厌氧条件下,SOB加强向胞内吸收硫化物,相较于对照组,液相中的硫化物浓度减少了50%,进而抑制了硫代硫酸盐的化学生成。除此之外,Kiragosyan等(2020)[75]研究发现,二甲基二硫化物可有效抑制硫酸盐生成,并通过向生物脱硫反应系统中添加二甲基二硫化物的方式,使生物源单质硫得率由92%进一步提升至96%。
3.2 单质硫的分离与回收为了实现生物源单质硫资源的回收与利用,研究者们根据生物源单质硫的组成和结构特性,采用物理方法从脱硫废水和脱硫污泥中回收生物源单质硫。常用的分离方法有3种:重力沉降法、离心分离法和过滤法。
由于单质硫在水中的溶解度很小,可以利用密度差,通过重力沉降法将单质硫从生物脱硫系统中分离出来[77-78]。与其他分离技术相比,重力沉降法是最经济、最具有吸引力的方法,单质硫分离效率可达到99.2%。然而,形成易于沉降的单质硫是一个重要的先决条件。生物源单质硫通常以胶体的形式存在,反应系统出水往往呈现乳白色,不能直接采用重力沉降法加以分离[8]。因此,有必要预先对反应系统出水采用酸处理,使生物源单质硫失稳,再用斜板沉淀法回收[79]。因经酸处理、斜板沉淀得到的浓缩液中单质硫含量为3-5 g/L,含水率为99%左右,需要进一步降低含水率[6]。除此之外,重力沉降法所需的沉降时间偏长,也需优化。
离心分离法是一种比较成熟的方法,已被广泛应用,比如,荷兰Paques公司研发的ThiopaqTM工艺中通过连续式离心机分离回收单质硫[5]。在回收单质硫之前,好氧生物反应器中产生的单质硫以10%固体含量的硫磺浆形式从液相分离出来[5]。用连续倾析式离心机对硫磺浆进行脱水,得到60%-65%干固体含量的硫磺饼,离心水作为工艺补充水返回生物反应器。离心后,单质硫纯度为95%-98%,其余2%-5%为无机盐,主要成分为碳酸氢钠和硫酸钠[6]。若进一步用于工业或农业,还需对硫磺进行洗涤、干燥和提炼,使其纯度达到99.9%以上[80]。
Zhang等(2006)[81]采用表面过滤法回收单质硫,表面过滤对过滤压力要求较低(压差大于7.0×105 Pa),更容易分离和回收过滤膜表面的单质硫。通过改变过滤膜内的流体力学条件,比如增加膜内流量,可将附着在膜上的单质硫高效分离[82]。然而,因硫化物氧化为单质硫是致碱反应,将加剧膜表面碳酸钙的沉积,堵塞过滤膜,降低单质硫的分离效率,因此对单质硫分离过程中的膜污染应深入研究[6]。
4 总结和展望单质硫是还原态含硫化合物生物氧化产生的中间产物。在SOB中,它既可贮存于胞内,也可排放到胞外。从生物脱硫系统中回收单质硫可实现硫循环利用。本文综述了SOB源单质硫的生成过程、赋存形式、代谢特性、转运机理、生物脱硫系统中单质硫回收等方面的研究进展。但这一领域仍有诸多盲点,建议今后加强以下方面的研究。
(1) 关于硫磺细菌资源,菌种是生物源单质硫回收的基础,获得高效泌硫型硫磺细菌,便可创建高效生物源单质硫回收技术。在今后的工作中,可开展从硫化细菌中筛选硫磺细菌,再从硫磺细菌中筛选泌硫型硫磺细菌的研究。
(2) 关于单质硫分泌机理,泌硫是生物源单质硫回收的推手,明晰泌硫机理,便可优化生物源单质硫回收技术。在今后的工作中,可开展单质硫赋存形态,单质硫运输载体,单质硫转运方式的研究。
(3) 关于单质硫得率,得率是生物源单质硫回收的关键,弄清单质硫得率的影响因素,便可强化生物源单质硫回收技术。在今后的工作中,可开展基质(硫化物和氧气)浓度,基质计量比,基质供给方式等因素对单质硫选择率影响的研究。
| [1] | Pokorna D, Zabranska J. Sulfur-oxidizing bacteria in environmental technology. Biotechnology Advances, 2015, 33(6): 1246-1259. DOI:10.1016/j.biotechadv.2015.02.007 |
| [2] | Maki JS. Bacterial intracellular sulfur globules: structure and function. Journal of Molecular Microbiology and Biotechnology, 2013, 23(4/5): 270-280. |
| [3] | Dahl C, Prange A. Bacterial sulfur globules: occurrence, structure and metabolism//Shively JM. Inclusions in Prokaryotes. Berlin, Heidelberg: Springer, 2006. |
| [4] | Kleinjan WE, de Keizer A, Janssen AJH. Biologically produced sulfur//Steudel R. Elemental Sulfur and Sulfur-Rich Compounds I. Berlin, Heidelberg: Springer, 2003. |
| [5] | Cline C, Hoksberg A, Abry R, Janssen A. Biological process for H2S removal from gas streams the shell-Paques/THIOPAQ™ gas desulfurization process//Proceedings of 2003 LRGCC. Norman, USA, 2003. |
| [6] | Cai J, Zheng P, Qaisar M, Zhang JQ. Elemental sulfur recovery of biological sulfide removal process from wastewater: a review. Critical Reviews in Environmental Science and Technology, 2017, 47(21): 2079-2099. DOI:10.1080/10643389.2017.1394154 |
| [7] | Xu GH, Yin FJ, Chen SH, Xu YJ, Yu HQ. Mathematical modeling of autotrophic denitrification (AD) process with sulphide as electron donor. Water Research, 2016, 91: 225-234. DOI:10.1016/j.watres.2016.01.011 |
| [8] | Li WJ, Zhang M, Kang D, Chen WD, Yu T, Xu DD, Zeng Z, Li YY, Zheng P. Mechanisms of sulfur selection and sulfur secretion in a biological sulfide removal (BISURE) system. Environment International, 2020, 137: 105549. DOI:10.1016/j.envint.2020.105549 |
| [9] | Cui YX, Guo G, Ekama GA, Deng YF, Chui HK, Chen GH, Wu D. Elucidating the biofilm properties and biokinetics of a sulfur-oxidizing moving-bed biofilm for mainstream nitrogen removal. Water Research, 2019, 162: 246-257. DOI:10.1016/j.watres.2019.02.061 |
| [10] | de Rink R, Klok JBM, van Heeringen GJ, Sorokin DY, ter Heijne A, Zeijlmaker R, Mos YM, de Wilde V, Keesman KJ, Buisman CJN. Increasing the selectivity for sulfur formation in biological gas desulfurization. Environmental Science & Technology, 2019, 53(8): 4519-4527. |
| [11] | Kiragosyan K, Klok JBM, Keesman KJ, Roman P, Janssen AJH. Development and validation of a physiologically based kinetic model for starting up and operation of the biological gas desulfurization process under haloalkaline conditions. Water Research X, 2019, 4: 100035. DOI:10.1016/j.wroa.2019.100035 |
| [12] | ter Heijne A, de Rink R, Liu DD, Klok JBM, Buisman CJN. Bacteria as an electron shuttle for sulfide oxidation. Environmental Science & Technology Letters, 2018, 5(8): 495-499. |
| [13] | Gregersen LH, Bryant DA, Frigaard NU. Mechanisms and evolution of oxidative sulfur metabolism in green sulfur bacteria. Frontiers in Microbiology, 2011, 2: 116. |
| [14] |
Liu Y, Jiang LJ, Shao ZZ. Advances in sulfur-oxidizing bacterial taxa and their sulfur oxidation pathways. Acta Microbiologica Sinica, 2018, 58(2): 191-201.
(in Chinese) 刘阳, 姜丽晶, 邵宗泽. 硫氧化细菌的种类及硫氧化途径的研究进展. 微生物学报, 2018, 58(2): 191-201. |
| [15] | Griesbeck C, Hauska G, Schütz M. Biological sulfide oxidation: Sulfide-Quinone Reductase (SQR), the primary reaction//Pandalai SG. Recent Research Developments in Microbiology. Trivadrum, India, 2000: 179-203. |
| [16] | Mitchell P, Moyle J. Chemiosmotic hypothesis of oxidative phosphorylation. Nature, 1967, 213(5072): 137-139. DOI:10.1038/213137a0 |
| [17] | Collman JP, Ghosh S, Dey A, Decréau RA. Using a functional enzyme model to understand the chemistry behind hydrogen sulfide induced hibernation. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(52): 22090-22095. DOI:10.1073/pnas.0904082106 |
| [18] | Klok JBM, de Graaff M, van den Bosch PLF, Boelee NC, Keesman KJ, Janssen AJH. A physiologically based kinetic model for bacterial sulfide oxidation. Water Research, 2013, 47(2): 483-492. |
| [19] | Brune DC. Sulfur oxidation by phototrophic bacteria. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 1989, 975(2): 189-221. DOI:10.1016/S0005-2728(89)80251-8 |
| [20] | Muyzer G, Sorokin DY, Mavromatis K, Lapidus A, Clum A, Ivanova N, Pati A, d'Haeseleer P, Woyke T, Kyrpides NC. Complete genome sequence of "Thioalkalivibrio sulfidophilus" HL-EbGr7. Standards in Genomic Sciences, 2011, 4(1): 23-35. DOI:10.4056/sigs.1483693 |
| [21] | Grimm F, Dobler N, Dahl C. Regulation of dsr genes encoding proteins responsible for the oxidation of stored sulfur in Allochromatium vinosum. Microbiology, 2010, 156(3): 764-773. |
| [22] | Müller AL, Kjeldsen KU, Rattei T, Pester M, Loy A. Phylogenetic and environmental diversity of DsrAB-type dissimilatory (bi) sulfite reductases. The ISME Journal, 2015, 9(5): 1152-1165. DOI:10.1038/ismej.2014.208 |
| [23] | Lin S, Mackey HR, Hao TW, Guo G, van Loosdrecht MCM, Chen GH. Biological sulfur oxidation in wastewater treatment: a review of emerging opportunities. Water Research, 2018, 143: 399-415. DOI:10.1016/j.watres.2018.06.051 |
| [24] | Cui YX, Biswal BK, Guo G, Deng YF, Huang H, Chen GH, Wu D. Biological nitrogen removal from wastewater using sulphur-driven autotrophic denitrification. Applied Microbiology and Biotechnology, 2019, 103(15): 6023-6039. DOI:10.1007/s00253-019-09935-4 |
| [25] | Frigaard NU, Dahl C. Sulfur metabolism in phototrophic sulfur bacteria. Advances in Microbial Physiology, 2008, 54: 103-200. DOI:10.1016/S0065-2911(08)00002-7 |
| [26] | Verté F, Kostanjevecki V, de Smet L, Meyer TE, Cusanovich MA, van Beeumen JJ. Identification of a thiosulfate utilization gene cluster from the green phototrophic bacterium Chlorobium limicola. Biochemistry, 2002, 41(9): 2932-2945. DOI:10.1021/bi011404m |
| [27] | Xia YZ, Lü CJ, Hou NK, Xin YF, Liu JH, Liu HL, Xun LY. Sulfide production and oxidation by heterotrophic bacteria under aerobic conditions. The ISME Journal, 2017, 11(12): 2754-2766. DOI:10.1038/ismej.2017.125 |
| [28] | Lü CJ, Xia YZ, Liu DX, Zhao R, Gao R, Liu HL, Xun LY. Cupriavidus necator H16 uses flavocytochrome c sulfide dehydrogenase to oxidize self-produced and added sulfide. Applied and Environmental Microbiology, 2017, 83(22): e01610-17. |
| [29] | Zhang YF, Weiner JH. Characterization of the kinetics and electron paramagnetic resonance spectroscopic properties of Acidithiobacillus ferrooxidans sulfide: quinone oxidoreductase (SQR). Archives of Biochemistry and Biophysics, 2014, 564: 110-119. DOI:10.1016/j.abb.2014.09.016 |
| [30] | Wang R, Lin JQ, Liu XM, Pang X, Zhang CJ, Yang CL, Gao XY, Lin CM, Li YQ, Li Y, Lin JQ, Chen LX. Sulfur oxidation in the acidophilic autotrophic Acidithiobacillus spp.. Frontiers in Microbiology, 2019, 9: 3290. DOI:10.3389/fmicb.2018.03290 |
| [31] | Cherney MM, Zhang YF, Solomonson M, Weiner JH, James MNG. Crystal structure of sulfide: Quinone oxidoreductase from Acidithiobacillus ferrooxidans: insights into sulfidotrophic respiration and detoxification. Journal of Molecular Biology, 2010, 398(2): 292-305. |
| [32] | Steudel R. Mechanism for the formation of elemental sulfur from aqueous sulfide in chemical and microbiological desulfurization processes. Industrial & Engineering Chemistry Research, 1996, 35(4): 1417-1423. |
| [33] | Schmidt GL, Nicolson GL, Kamen MD. Composition of the sulfur particle of Chromatium vinosum strain D. Journal of Bacteriology, 1971, 105(3): 1137-1141. DOI:10.1128/JB.105.3.1137-1141.1971 |
| [34] | Clayton RK, Sistrom WR. The photosynthetic bacteria. New York: Plenum Press, 1978. |
| [35] | Schmidt TM, Vinci VA, Strohl WR. Protein synthesis by Beggiatoa alba B18LD in the presence and absence of sulfide. Archives of Microbiology, 1986, 144(2): 158-162. DOI:10.1007/BF00414727 |
| [36] | Brune DC. Isolation and characterization of sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina. Archives of Microbiology, 1995, 163(6): 391-399. DOI:10.1007/BF00272127 |
| [37] | Pattaragulwanit K, Brune DC, Trüper HG, Dahl C. Molecular genetic evidence for extracytoplasmic localization of sulfur globules in Chromatium vinosum. Archives of Microbiology, 1998, 169(5): 434-444. DOI:10.1007/s002030050594 |
| [38] | Prange A, Engelhardt H, Trüper HG, Dahl C. The role of the sulfur globule proteins of Allochromatium vinosum: mutagenesis of the sulfur globule protein genes and expression studies by real-time RT-PCR. Archives of Microbiology, 2004, 182(2/3): 165-174. DOI:10.1007/s00203-004-0683-3 |
| [39] | Lawry NH, Jani V, Jensen TE. Identification of the sulfur inclusion body in Beggiatoa alba B18LD by energy-dispersive X-ray microanalysis. Current Microbiology, 1981, 6(2): 71-74. DOI:10.1007/BF01569006 |
| [40] | Prange A, Arzberger I, Engemann C, Modrow H, Schumann O, Trüper HG, Steudel R, Dahl C, Hormes J. In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteria by X-ray absorption near edge spectroscopy. Biochimica et Biophysica Acta (BBA)-General Subjects, 1999, 1428(2/3): 446-454. |
| [41] | Prange A, Chauvistré R, Modrow H, Hormes J, Trüper HG, Dahl C. Quantitative speciation of sulfur in bacterial sulfur globules: X-ray absorption spectroscopy reveals at least three different species of sulfur. Microbiology, 2002, 148(1): 267-276. |
| [42] | Teske A, Nelson DC. The genera Beggiatoa and Thioploca//Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E. The Prokaryotes: Volume 6: Proteobacteria: Gamma Subclass. New York: Springer, 2006: 784-810. |
| [43] | Strohl WR, Larkin JM. Enumeration, isolation, and characterization of Beggiatoa from freshwater sediments. Applied and Environmental Microbiology, 1978, 36(5): 755-770. |
| [44] | Strohl WR, Geffers I, Larkin JM. Structure of the sulfur inclusion envelopes from four beggiatoas. Current Microbiology, 1981, 6(2): 75-79. DOI:10.1007/BF01569007 |
| [45] | Janssen A, de Keizer A, van Aelst A, Fokkink R, Yangling H, Lettinga G. Surface characteristics and aggregation of microbiologically produced sulphur particles in relation to the process conditions. Colloids and Surfaces B: Biointerfaces, 1996, 6(2): 115-129. |
| [46] | Lee YJ, Prange A, Lichtenberg H, Rohde M, Dashti M, Wiegel J. In situ analysis of sulfur species in sulfur globules produced from thiosulfate by Thermoanaerobacter sulfurigignens and Thermoanaerobacterium thermosulfurigenes. Journal of Bacteriology, 2007, 189(20): 7525-7529. |
| [47] | Sowerby DB, Haiduc I. The chemistry of inorganic homo-and hetero-cycles. London: Academic, 1987. |
| [48] | Boulegue J. Solubility of elemental sulfur in water at 298 K. Phosphorus and Sulfur and the Related Elements, 1978, 5(1): 127-128. |
| [49] | Jones GE, Starkey RL. Surface-active substances produced by Thiobacillus thiooxidans. Journal of Bacteriology, 1961, 82(5): 788-789. |
| [50] | Jones GE, Benson AA. Phosphatidyl glycerol in Thiobacillus thiooxidans. Journal of Bacteriology, 1965, 89(1): 260-261. |
| [51] | Marnocha CL, Levy AT, Powell DH, Hanson TE, Chan CS. Mechanisms of extracellular S0 globule production and degradation in Chlorobaculum tepidum via dynamic cell-globule interactions. Microbiology, 2016, 162(7): 1125-1134. |
| [52] | Hanson TE, Bonsu E, Tuerk A, Marnocha CL, Powell DH, Chan CS. Chlorobaculum tepidum growth on biogenic S(0) as the sole photosynthetic electron donor. Environmental Microbiology, 2016, 18(9): 2856-2867. |
| [53] | Cosmidis J, Nims CW, Diercks D, Templeton AS. Formation and stabilization of elemental sulfur through organomineralization. Geochimica et Cosmochimica Acta, 2019, 247: 59-82. |
| [54] | Cron B, Henri P, Chan CS, Macalady J, Cosmidis J. Elemental sulfur formation by Sulfuricurvum kujiense is mediated by extracellular organic compounds. Frontiers in Microbiology, 2019, 10: 2710. |
| [55] | Rickard D, Luther GW. Chemistry of iron sulfides. Chemical Reviews, 2007, 107(2): 514-562. |
| [56] | Marnocha CL, Sabanayagam CR, Modla S, Powell DH, Henri PA, Steele AS, Hanson TE, Webb SM, Chan CS. Insights into the mineralogy and surface chemistry of extracellular biogenic S0 globules produced by Chlorobaculum tepidum. Frontiers in Microbiology, 2019, 10: 271. |
| [57] | Franz B, Lichtenberg H, Hormes J, Modrow H, Dahl C, Prange A. Utilization of solid 'elemental' sulfur by the phototrophic purple sulfur bacterium Allochromatium vinosum: a sulfur K-edge X-ray absorption spectroscopy study. Microbiology, 2007, 153(4): 1268-1274. |
| [58] | Beard S, Paradela A, Albar JP, Jerez CA. Growth of Acidithiobacillus ferrooxidans ATCC 23270 in thiosulfate under oxygen-limiting conditions generates extracellular sulfur globules by means of a secreted tetrathionate hydrolase. Frontiers in Microbiology, 2011, 2: 79. |
| [59] | Steudel R, Holdt G, Göbel DCT, Hazeu IW. Chromatographic separation of higher polythionates SnO62Θ (n=3…22) and their detection in cultures of Thiobacillus ferroxidans; molecular composition of bacterial Sulfur secretions. Angewandte Chemie International Edition in English, 1987, 26(2): 151-153. |
| [60] | Schwechheimer C, Kuehn MJ. Outer-membrane vesicles from Gram-negative bacteria: biogenesis and functions. Nature Reviews Microbiology, 2015, 13(10): 605-619. |
| [61] | Kulp A, Kuehn MJ. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annual Review of Microbiology, 2010, 64: 163-184. |
| [62] | MacDonald IA, Kuehn MJ. Stress-induced outer membrane vesicle production by Pseudomonas aeruginosa. Journal of Bacteriology, 2013, 195(13): 2971-2981. |
| [63] | McBroom AJ, Kuehn MJ. Release of outer membrane vesicles by Gram‐negative bacteria is a novel envelope stress response. Molecular Microbiology, 2007, 63(2): 545-558. |
| [64] | de Rink R, Klok JBM, van Heeringen GJ, Keesman KJ, Janssen AJH, ter Heijne A, Buisman CJN. Biologically enhanced hydrogen sulfide absorption from sour gas under haloalkaline conditions. Journal of Hazardous Materials, 2020, 383: 121104. |
| [65] | Janssen AJH, Sleyster R, van der Kaa C, Jochemsen A, Bontsema J, Lettinga G. Biological sulphide oxidation in a fed‐batch reactor. Biotechnology and Bioengineering, 1995, 47(3): 327-333. |
| [66] | van den Bosch PLF, van Beusekom OC, Buisman CJN, Janssen AJH. Sulfide oxidation at halo-alkaline conditions in a fed-batch bioreactor. Biotechnology and Bioengineering, 2007, 97(5): 1053-1063. |
| [67] | Klok JBM, van den Bosch PLF, Buisman CJN, Stams AJM, Keesman KJ, Janssen AJH. Pathways of sulfide oxidation by haloalkaliphilic bacteria in limited-oxygen gas lift bioreactors. Environmental Science & Technology, 2012, 46(14): 7581-7586. |
| [68] | Madigan M, Martinko J, Bender K, Buckley D, Stahl D. Brock Biology of Microorganisms. 14th ed. Glenview, IL, USA: Pearson, 2014. |
| [69] | Buisman CJN, Geraats BG, Ijspeert P, Lettinga G. Optimization of sulphur production in a biotechnological sulphide‐removing reactor. Biotechnology and Bioengineering, 1990, 35(1): 50-56. |
| [70] | Janssen AJH, Ma SC, Lens P, Lettinga G. Performance of a sulfide-oxidizing expanded-bed reactor supplied with dissolved oxygen. Biotechnology and Bioengineering, 1997, 53(1): 32-40. |
| [71] | Lohwacharin J, Annachhatre AP. Biological sulfide oxidation in an airlift bioreactor. Bioresource Technology, 2010, 101(7): 2114-2120. |
| [72] | Janssen AJH, Meijer S, Bontsema J, Lettinga G. Application of the redox potential for controling a sulfide oxidizing bioreactor. Biotechnology and Bioengineering, 1998, 60(2): 147-155. |
| [73] | Krishnakumar B, Majumdar S, Manilal VB, Haridas A. Treatment of sulphide containing wastewater with sulphur recovery in a novel reverse fluidized loop reactor (RFLR). Water Research, 2005, 39(4): 639-647. |
| [74] | Alcántara S, Velasco A, Muñoz A, Cid J, Revah S, Razo-Flores E. Hydrogen sulfide oxidation by a microbial consortium in a recirculation reactor system: sulfur formation under oxygen limitation and removal of phenols. Environmental Science & Technology, 2004, 38(3): 918-923. |
| [75] | Kiragosyan K, Picard M, Sorokin DY, Dijkstra J, Klok JBM, Roman P, Janssen AJH. Effect of dimethyl disulfide on the sulfur formation and microbial community composition during the biological H2S removal from sour gas streams. Journal of Hazardous Materials, 2020, 386: 121916. |
| [76] | van den Bosch PLF. Biological sulfide oxidation by natron-alkaliphilic bacteria: application in gas desulfurization. Doctor Dissertation of Wageningen University, 2008. |
| [77] | Berg JS, Schwedt A, Kreutzmann AC, Kuypers MMM, Milucka J. Polysulfides as intermediates in the oxidation of sulfide to sulfate by Beggiatoa spp.. Applied and Environmental Microbiology, 2014, 80(2): 629-636. |
| [78] | Muyzer G, Kuenen JG, Robertson LA. Colorless sulfur bacteria//Rosenberg E, DeLong EF, Lory S, Stackebrandt E, Thompson F. The Prokaryotes: Prokaryotic Physiology and Biochemistry. Berlin, Heidelberg: Springer, 2013: 555-588. |
| [79] | Chen F, Yuan Y, Chen C, Zhao YK, Tan WB, Huang C, Xu XJ, Wang AJ. Investigation of colloidal biogenic sulfur flocculation: Optimization using response surface analysis. Journal of Environmental Sciences, 2016, 42: 227-235. |
| [80] | Janssen AJH, Ruitenberg R, Buisman CJN. Industrial applications of new sulphur biotechnology. Water Science and Technology, 2001, 44(8): 85-90. |
| [81] |
Zhang KQ, Huang WX, Ji M, Li JX, Li Y, Li B, Zhang JF. Characteristics of microbiologically sulfur producing during biological treatment for wastewater containing sulfide. Journal of Agro-Environment Science, 2006, 25(2): 522-526.
(in Chinese) 张克强, 黄文星, 季民, 李军幸, 李野, 李斌, 张金凤. 含硫化物废水生物处理过程中单质硫的形成特性. 农业环境科学学报, 2006, 25(2): 522-526. |
| [82] | Sahinkaya E, Hasar H, Kaksonen AH, Rittmann BE. Performance of a sulfide-oxidizing, sulfur-producing membrane biofilm reactor treating sulfide-containing bioreactor effluent. Environmental Science & Technology, 2011, 45(9): 4080-4087. |