 2022, Vol. 62
2022, Vol. 62
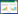


NLRP6在感染性和非感染性疾病中的作用

http://dx.doi.org/10.13343/j.cnki.wsxb.20210452
中国科学院微生物研究所,中国微生物学会
中国科学院微生物研究所,中国微生物学会
文章信息
- 陶旗, 徐冬怡, 方仁东. 2022
- TAO Qi, XU Dongyi, FANG Rendong.
- NLRP6在感染性和非感染性疾病中的作用
- Role of NLRP6 in infectious and non-infectious diseases
- 微生物学报, 62(4): 1190-1201
- Acta Microbiologica Sinica, 62(4): 1190-1201
-
文章历史
- 收稿日期:2021-07-30
- 修回日期:2021-09-16
- 网络出版日期:2021-09-24
引用本文 |
陶旗, 徐冬怡, 方仁东. NLRP6在感染性和非感染性疾病中的作用[J]. 微生物学报, 2022, 62(4): 1190-1201.
Tao Qi, Xu Dongyi, Fang Rendong. Role of NLRP6 in infectious and non-infectious diseases[J]. Acta Microbiologica Sinica, 2022, 62(4): 1190-1201.
NLRP6在感染性和非感染性疾病中的作用
1. 西南大学动物医学院动物健康与动物性食品安全国际合作联合实验室, 重庆 400715;
2. 西南大学医学研究院免疫学研究中心, 重庆 402460;
3. 草食动物科学重庆市重点实验室, 重庆 400715
2. 西南大学医学研究院免疫学研究中心, 重庆 402460;
3. 草食动物科学重庆市重点实验室, 重庆 400715
摘要:核苷酸结合寡聚化结构域样受体含pyrin结构域蛋白6 (NLRP6)属于核苷酸结合寡聚化结构域样受体(NLRs)家族蛋白。NLRP6可以作为受体蛋白参与炎症小体的组装,引起白细胞介素IL-1β和IL-18的成熟与分泌。NLRP6在病原体感染诱导宿主天然免疫应答中发挥重要作用。此外,NLRP6在维持肠道上皮完整、调控代谢性疾病及神经性炎症等方面也发挥重要功能。本文将对NLRP6炎症小体的激活与调控机制、NLRP6在感染性和非感染性疾病中的作用等研究进展进行综述。
关键词:NLRP6 炎症小体 宿主防御
Role of NLRP6 in infectious and non-infectious diseases
1. Joint International Research Laboratory of Animal Health and Animal Food Safety, College of Veterinary Medicine, Southwest University, Chongqing 400715, China;
2. Immunology Research Center, Medical Research Institute, Southwest University, Chongqing 402460, China;
3. Chongqing Key Laboratory of Herbivore Science, Chongqing 400715, China
2. Immunology Research Center, Medical Research Institute, Southwest University, Chongqing 402460, China;
3. Chongqing Key Laboratory of Herbivore Science, Chongqing 400715, China
Abstract: Nucleotide-binding oligomerization domain (NOD)-like receptor family pyrin domain containing 6 (NLRP6) is a member of the NOD-like receptor (NLR) family. It participates in inflammasome assembly to mediate the maturation and secretion of the pro-inflammatory cytokines IL-1β and IL-18. In addition, it plays an important role in host innate immune response to pathogen, maintaining intestinal epithelial integrity, and regulating metabolic diseases and neuroinflammation. In this paper, we reviewed the activation and regulation mechanism of NLRP6 inflammasome, as well as the role of NLRP6 in infectious and non-infectious diseases.
Keywords:
NLRP6 inflammasome host defense
先天免疫系统作为宿主防御的第一道防线,依赖于模式识别受体(pattern recognition receptors,PRRs)识别病原体相关分子模式(pathogen-associated molecular patterns,PAMPs)和损伤相关分子模式(damage-associated molecular patterns,DAMPs)产生免疫反应[1–2]。PRRs包括Toll样受体(Toll-like receptors,TLRs)和NOD样受体(NOD-like receptors,NLRs)等,PRRs介导的信号传导级联反应会引发一系列先天免疫应答,包括炎症小体的激活、细胞因子分泌和细胞焦亡[3–4]。NLRP6最早由Grenier等[5]于2002年提出,与家族中其他成员一样,由C端富含亮氨酸的重复序列(leucine-rich repeats,LRR)、N端热蛋白结构域(pyrin domain,PYD)以及中间核苷酸偶联结构域(NACHT/ NOD) 3部分组成,LRR能识别不同的DAMPs和PAMPs[6–7]。NLRP6在骨髓源细胞中表达,包括树突状细胞、中性粒细胞、巨噬细胞和T细胞,也在肠道上皮细胞中高表达[8–11]。NLRP6可以组装成炎症小体复合物介导促炎细胞因子IL-1β、IL-18成熟分泌,并且可以裂解消皮素D (GSDMD)诱导细胞焦亡,也可独立于炎症小体发挥功能[12–13]。NLRP6已被证明在调节炎症和宿主防御中发挥重要作用,本文结合本课题组及国内外最新研究进展,就NLRP6炎症小体的激活机制、NLRP6在感染性与非感染性疾病中的作用等方面进行阐述。
1 NLRP6炎症小体激活与调控机制 1.1 NLRP6炎症小体的组装与激活炎症小体是胞浆内一种蛋白复合体,通过募集半胱天冬酶(caspases)来促进细胞因子成熟与分泌,诱导细胞焦亡,在识别病原体并产生免疫应答中起关键作用[14]。NLRPs是NLRs家族中最大的蛋白亚族,NLRP6在某些条件下被激活后通过PYD-PYD同源蛋白相互作用招募凋亡相关斑点样蛋白(apoptosis-associated speck-like protein containing a CARD,ASC),ASC进一步通过半胱天冬酶募集结构域CARD与caspase-1和/或caspase-11结合形成完整的NLRP6炎症小体[13, 15–16]。最近,Shen等[17]通过冷冻电镜技术和分子动力学方法揭示了NLRP6的PYD结构域能够自组装成丝状结构,伴随构象变化,募集ASC的PYD结构域。
越来越多的数据表明,多种微生物组分和代谢物可以调控NLRP6炎症小体组装。微生物代谢产物牛磺酸可以诱导NLRP6炎症小体激活,促进IL-18成熟分泌,IL-18又能够调节抗菌肽产生,在肠道中形成化学屏障,保护肠道免受炎症损伤,组胺和精胺则表现出抑制作用[18]。另有研究表明,胃旁路术(Roux-en-Y gastric bypass)通过提高肠道中牛磺酸水平来活化NLRP6炎症小体,从而恢复肠道通透性,减轻炎症[19]。
除微生物代谢物与NLRP6炎症小体激活有关,单增李斯特菌(Listeria monocytogens)细胞壁组分脂磷壁酸(lipoteichoic acid,LTA)可以结合并激活NLRP6炎症小体[20]。对于革兰阴性菌,细胞外膜主要成分脂多糖(lipopolysaccharide,LPS)可直接与NLRP6结合,诱导NLRP6整体构象改变形成二聚体,在ATP的刺激下二聚体组装成四聚体、六聚体甚至十二聚体等低聚物以此来招募ASC[21]。Lu等[22]使用大肠杆菌(Escherichia coli) LPS刺激人牙周膜细胞后,NLRP6表达上调。此外,去泛素化酶Cyld可抑制NLRP6与ASC结合,减少IL-18成熟,防止过度炎症[23]。
1.2 Nlrp6转录调控Nlrp6基因转录表达受到多种物质的调控,Nlrp6基因启动子区存在大量的过氧化物酶体增殖物激活受体(peroxisome proliferators-activated receptor γ,PPARγ)、视黄醇受体α (retinoid X receptor α,RXRα)和鸡卵清蛋白上游启动子转录因子1 (chicken ovalbumin upstream promoter transcription factor 1,COUP-TF 1)结合序列[24]。人上皮细胞系Caco-2与PPARγ激动剂罗格列酮共培养后,Nlrp6 mRNA表达上调,并且罗格列酮能够诱导肠上皮细胞表达NLRP6,减轻避水应激(water-avoidance stress)引起的肠炎。以上体内外试验表明PPARγ参与Nlrp6的代谢调节[24–25]。同样利用Caco-2细胞系,无论有无活性的白色念珠菌(Candida albicans)均能够抑制Nlrp6 mRNA转录[26]。最近研究发现,Nlrp6 mRNA转录水平受到表观遗传学调控,microRNA-650 (miR-650)作为Nlrp6基因的上游调控因子,在溃疡性结肠炎的发病机制中发挥重要作用,miR-650通过与Nlrp6的3′端非编码区结合抑制NLRP6表达[27]。在脑内出血(intracerebral hemorrhage,ICH)的研究中发现microRNA-331-3p (miR-331-3p)也能负调控NLRP6的表达[28]。NLRP6之前被证实可以抑制胃肿瘤的发生,长链非编码RNA OIP5-AS1(lncRNA OIP5-AS1)可以直接与组蛋白甲基化转移酶2 (enhancer of zeste homolog 2,EZH2)结合抑制NLRP6表达,进而促进胃癌发生[29–30]。NLRP6表达还受到激素调节,促肾上腺皮质激素在肠易激综合征(irritable bowel syndrome)中释放增加并下调NLRP6表达[25, 31]。
1.3 NLRP6对NF-κB和MAPK信号通路的调控与其他NOD样受体不同,NLRP6不仅通过产生IL-1β和IL-18参与炎症反应,还可以抑制核因子κB (nuclear factor kappa-B,NF-κB)和丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号通路从而抑制炎症和预防病理损伤。NLRP6敲除(NLRP6–/–)小鼠巨噬细胞感染单增李斯特菌后或在暴露于TLR2/4配体时,造成TNF-α和IL-6等细胞因子表达上升,表明NLRP6特异性抑制TLR2/4依赖的NF-κB和MAPK通路激活[32]。在酒精性肝炎(alcoholic hepatitis,AH)和非酒精性脂肪性肝炎(non-alcoholic steatohepatitis,NASH)模型中,NLRP6均能抑制NF-κB信号通路的激活,起到保护肝脏作用[33–34]。本课题组最新研究发现,在肺炎链球菌(Streptococcus pneumoniae)分别感染NLRP6–/–、WT小鼠的腹腔巨噬细胞时,NLRP6敲除组中的p-p65、p-ERK1/2和p-IκBα表达水平明显升高[35],表明NLRP6负调节炎症性NF-κB和ERK信号通路。NLRP6缺失时,NLRP3激活可能作为一种补偿机制,两种炎症小体在多数情况下表现出相反作用,但是这两种炎症小体之间具体是如何调控的尚不清楚,需更多实验数据加以阐述。鼠NLRP6和NLRP3氨基酸序列相似度高达33%,主要差异位于LRR结构域[36],提示NLRP6与NLRP3能识别不同配体。将Nlrp6基因沉默,大肠杆菌LPS刺激人牙周膜细胞可诱导NF-κB和ERK信号通路激活,IL-6和TNF-α水平升高,同时NLRP3、pro-IL-1β和pro-caspase-1表达上调[22]。Chen等[37]发现在碱烧伤后,caspase-8信号通路通过NF-κB途径上调NLRP3表达,而NLRP6表达则被抑制。在异基因造血干细胞移植诱导的肝损伤中,NLRP6–/–小鼠相对于野生型(WT)小鼠,IκBα、p38-MAPK磷酸化增强,以及细胞因子IL-1β、IL-18、IL-6和TNF-α表达增加,NLRP3表达水平也明显升高[38]。
2 NLRP6与感染性疾病NLR家族成员比如NOD1、NOD2、NLRC4以及NLRP3对清除微生物病原体起到积极保护作用,缺失这些受体将增加宿主的发病率和死亡率[39–42]。NLRP6作为NLR家族成员之一,对宿主防御细菌、病毒和寄生虫等病原体同样具有重要调节作用。在不同微生物感染肺脏和肠道期间,NLRP6表现出对宿主起保护或损害作用(表 1)。
表 1. NLRP6调节宿主防御病原微生物
Table 1. NLRP6 regulates host defense against pathogenic microorganisms
| Pathogens | Tissue | NLRP6 plays a protective or damaging role | Mechanism of action | References |
| S. aureus | Lung | Damage | Deletion of NLRP6 enhances bacterial killing of neutrophils through increased IFN-γ and ROS production. | [43] |
| K. pneumoniae | Lung | Protection | NLRP6 promotes NET formation and NET-mediated bacterial killing. | [44] |
| S. pneumoniae | Lung | Damage | NLRP6 inhibits the recruitment of neutrophils and macrophages in the lungs. | [35] |
| L. monocytogens | Intestine | Damage | Deletion of NLRP6 enhances the NF-κB and MAPK signaling pathways, and increases the recruitment of neutrophils, which is conducive to enhancing the ability of host bacteria to clear. NLRP6 inflammasome mediates the secretion of IL-18, which aggravates systemic bacterial infections. | [8, 20] |
| S. typhimurium | Intestine | Damage | NLRP6 inhibits neutrophil recruitment mediated by NF-κB signaling pathway. | [8] |
| E. coli | Intestine | Damage | Deletion of NLRP6 increases neutrophil recruitment. | [8] |
| C. rodentium | Intestine | Protection | NLRP6 regulates the secretion of mucus by goblet cells to form mucosal defense bacteria. | [46–47] |
| EMCV, norovirus | Intestine | Protection | The activated NLRP6-DHX15-MAVS axis augments the expression of ISGs to limit virus in the intestine. | [48] |
| C. tyzzeri | Intestine | Protection | NLRP6 inflammasome eliminates Cryptosporidium by promoting the maturation of IL-18 and GSDMD. | [50] |
2.1 NLRP6在病原体感染肺脏中的作用
Ghimire等[43]用金黄色葡萄球菌(Staphylococcus aureus)感染WT和NLRP6–/–小鼠的肺炎模型研究发现,与WT小鼠相比,NLRP6–/–小鼠对细菌的敏感性降低,表现出更高的存活率和细菌清除力;同时其肺部中性粒细胞、自然杀伤性细胞和CD4+T细胞募集增加。金黄色葡萄球菌(Staphylococcus aureus)作为细胞外病原菌,通过NLRP6诱导细胞坏死性凋亡,促进炎症发生,最终对宿主产生损害作用,但NLRP6调控细胞坏死性凋亡的具体机制还有待进一步研究。
在大多数研究中,NLRP6是作为负调控因子抑制宿主的防御应答,但是在肺炎克雷伯菌(Klebsiella pneumoniae)败血症中,NLRP6能够调节中性粒细胞的稳态。NLRP6–/–小鼠感染肺炎克雷伯菌后,小鼠肺泡灌洗液中中性粒细胞数量下降,髓过氧化物酶(myeloperoxidase,MPO)分泌减少,趋化因子(CXCL1、CXCL2和CXCL5)以及细胞因子(TNF-α、IL-6、G-CSF、IL-β)表达下降,细菌向脾脏扩散导致死亡率升高[44]。中性粒细胞外诱捕网(NETs)的形成对中性粒细胞杀死细胞外病原体至关重要[45],将Nlrp6基因敲除后,小鼠NETs形成受损,肺部细菌定殖量上升。进一步研究发现,向NLRP6–/–小鼠注射重组CXCL1后,宿主防御能力增强,NETs形成得到修复[44]。因此推测NLRP6作为中性粒细胞募集、生成和功能的中心调节因子,在肺炎克雷伯菌感染肺部期间起保护作用。
本课题组通过NLRP6–/–小鼠模型证实NLRP6在肺炎链球菌感染防御中起损害作用[35]。与WT小鼠相比,NLRP6–/–小鼠感染肺炎链球菌后,死亡率更低。WT小鼠肺脏细菌定殖量明显高于NLRP6–/–小鼠,肺部中性粒细胞与巨噬细胞募集降低,炎症加重,表现为间质增生、炎性细胞浸润等[35]。NLRP6在肺炎链球菌感染过程中的损害作用可能是由于其抑制肺部免疫细胞募集以及降低中性粒细胞细菌清除能力。
2.2 NLRP6在病原体感染肠道中的作用 2.2.1 NLRP6与肠道病原菌感染NLRP6在肠道中防御病原菌感染具有调节作用,Anand等[8]报道称NLRP6负调控炎症信号通路,NLRP6–/–小鼠对单增李斯特菌的耐药性增强。感染上述细菌后,NLRP6–/–小鼠各脏器细菌定殖量下降,具有更高的存活率,外周血中性粒细胞和单核细胞募集增加[8, 20]。NLRP6可能通过分泌IL-18加重单增李斯特菌全身感染,单增李斯特菌的LTA能够直接与NLRP6结合形成受体蛋白复合物,引起ASC寡聚化,同时募集caspase-1和caspase-11形成炎症小体,促进巨噬细胞分泌IL-18和IL-1β (图 1A),向敲除型小鼠注射IL-18后,小鼠对单增李斯特菌的易感性增加[20]。

|
| 图 1 NLRP6与肠道病原体相互作用 Figure 1 NLRP6 interacts with intestinal pathogens. A: LTA, a component of Listeria monocytogens, might bind directly to NLRP6, and then NLRP6 recruits ASC and pro-caspase-1/caspase-11 to form the NLRP6 inflammasome which cleaves pro-IL-18 into its active form. B: TLR ligands such as LPS, lipid A, and flagellin activate the Myd88-ROS pathway to activate the NLRP6 inflammasome, to facilitate exocytosis of mucin granules to form a colonic mucus layer above the epithelium. C: NLRP6 recognizes dsRNA of EMCV via the NLRP6–Dhx15 viral sensing complex. This complex further triggers the induction of type Ⅰ/Ⅲ IFNs and ISGs through MAVS. D: Cryptosporidium activates NLRP6 inflammasome leads to the processing and release of IL-18 in a GSDMD-dependent manner. |
为排除NLRP6调节宿主防御细菌病原体是与细菌特性有关,Anand等[8]还使用沙门氏菌(Salmonella typhimurium)和大肠杆菌(Escherichia coli)通过腹腔注射的感染方式感染WT小鼠和NLRP6–/–小鼠。结果与单增李斯特菌感染结果一致,NLRP6–/–小鼠的肝脏和脾脏细菌定殖量下降,宿主存活率上升[8]。
Wlodarska等[46]利用啮齿柠檬酸杆菌(Citrobacter rodentium)诱导的小鼠肠炎模型发现NLRP6通过调节杯状细胞分泌黏液维持肠道稳态。位于结肠隐窝处的杯状细胞能够非特异性地胞吞LPS、脂质A、三酰脂肽Pam3CSK4以及鞭毛蛋白等Toll样受体的配体,然后诱导胞内产生ROS,进一步激活NLRP6,随后调节杯状细胞分泌黏蛋白Muc2形成黏膜以防御细菌病原体[47](图 1B)。与WT小鼠相比,感染啮齿柠檬酸杆菌后,NLRP6–/–小鼠肠道细菌定殖量增加,黏膜溃疡、水肿和增生,宿主防御受损[46]。
2.2.2 NLRP6与肠道病毒感染NLRP6不仅能够调节抗菌免疫反应,Wang等[48]首次证实NLRP6通过炎症小体非依赖性途径在肠道的抗病毒反应中起关键作用。将脑心肌炎病毒(encephalomyocarditis virus,EMCV) 通过腹腔注射的方式感染WT小鼠和NLRP6–/–小鼠,发现两者的存活率相似,病毒量在血液、大脑以及心脏中没有明显差异,但是NLRP6–/–小鼠肠道中的病毒量明显高于WT小鼠。通过口服的感染方式感染EMCV,NLRP6–/–小鼠对病毒的易感性增强,死亡率升高。造成这一差异的原因可能是NLRP6在肠道中发挥抗病毒作用,肠道中的NLRP6能够抑制病毒全身扩散。鼠诺如病毒(Norovirus)感染宿主也得到类似的结果。该课题组进一步研究发现,NLRP6在宿主肠道上皮细胞中,可以和RNA解旋酶DHX15形成复合体识别并结合病毒RNA,进而激活线粒体抗病毒信号蛋白(MAVS)。激活的NLRP6-DHX15-MAVS轴促进干扰素刺激基因(ISGs)大量表达,从而达到抗病毒效果(图 1C)。该过程不依赖于经典炎症小体途径,且不改变肠道微生物组成。值得注意的是,NLRs家族中的另一成员NLRP9b在肠道中通过RNA解旋酶DHX9间接识别病毒RNA,并与ASC、caspase-1形成炎症小体促进IL-18和GSDMD成熟诱导细胞焦亡[49]。因此,NLRP6是否与其他炎症小体共同调节宿主对病毒的防御以及在其他脏器中防御病毒的具体机制还有待进一步研究。
2.2.3 NLRP6与肠道寄生虫感染最新研究证实,泰泽隐孢子虫(Cryptosporidium tyzzeri)感染肠道早期,NLRP6能够形成炎症小体防御其感染[50]。泰泽隐孢子虫在NLRP6–/–、ASC–/–、caspase-1–/–、GSDMD–/–和IL-18–/–小鼠肠道中的定殖量显著高于WT小鼠,向caspase1/11–/–小鼠注射重组IL-18后,小鼠对泰泽隐孢子虫易感性减弱[50]。IL-18是先天免疫控制隐孢子虫的重要媒介,故认为NLRP6炎症小体可能通过促进IL-18分泌来清除肠道的隐孢子虫(图 1D),但是这与单增李斯特菌感染肠道研究结果[20]相矛盾,因此阐明NLRP6识别并清除隐孢子的分子模式对了解肠细胞如何启动先天免疫应对肠道病原体具有重要意义。
总之,NLRP6在宿主防御不同病原体中发挥不同作用,NLRP6介导的宿主防御调控可能依赖于环境以及病原微生物本身。因此,还需要更多的研究来进一步定义NLRP6在先天免疫系统中的作用,挖掘NLRP6防御病原微生物的具体机制。
3 NLRP6与非感染性疾病炎症小体已被证明与肠道屏障完整性、微生物组成和肝脏损伤有关[51]。NLRP6在多种组织中均有表达,在小肠、大肠中表达量较高,脾脏和肺中表达程度较低[10–11]。NLRP6不仅与宿主防御多种病原体有关,近年来一些研究报道了NLRP6还参与肠道、肝脏等非感染性疾病的调控。
3.1 NLRP6与肠道疾病由于NLRP6在肠道中高表达,NLRP6功能的研究主要集中在肠道[52]。宿主免疫反应的功能障碍和肠道微生物群的病理改变都有助于炎症性肠病(inflammatory bowel disease,IBD)的发展[9]。NLRP6能够改变肠道微生物菌群组成结构,NLRP6缺乏会增加与IBD相关的菌群丰度,例如嗜黏蛋白-艾克曼菌、普氏菌和TM7细菌[53–54],但是目前尚不清楚这些细菌群的富集是否直接受NLRP6调控,需更进一步研究。利用硫酸葡聚糖钠盐(dextran sulfate sodium,DSS)诱导结肠炎发现NLRP6在Ly6Chi单核细胞中表达特异性上调,将WT小鼠Ly6Chi单核细胞转移到NLRP6–/–小鼠中,可显著降低肠道通透性和损伤,提高小鼠存活率[55]。另外,NLRP6表达降低可能导致巨结肠患者结肠微生物组成的改变,并促进巨结肠相关性小肠结肠炎(hirschsprung’s disease associated enterocolitis)发生[56]。由此可见,NLRP6能够保护小鼠免受化学诱导的肠道炎症,并与肠道微生物群的结构息息相关。
3.2 NLRP6与肝脏疾病NLRP6缺失会使TLR4激动剂流入门静脉循环导致肝脏脂肪变性和产生炎症[57]。异基因造血干细胞移植(allogeneic hematopoietic stem cell transplantation,allo-HSCT)是有效治疗血液疾病如白血病、多发性骨髓瘤、骨髓增生异常综合征和一些造血系统疾病的方法,但是,allo-HSCT会引起肝损伤等并发症,从而影响患者术后的生存率[58]。NLRP6在allo-HSCT后,肝组织中的表达升高,与WT小鼠相比,NLRP6–/–小鼠移植后的肝功能明显受损,表现为肝组织水肿加重,伴有炎性细胞浸润、血栓形成和纤维蛋白沉积增多,这说明NLRP6可以减轻allo-HSCT后的肝脏损伤,减少炎症细胞浸润和肝纤维化[38]。NLRP6还在酒精性肝炎发生发展中起保护作用,在患有酒精性肝炎小鼠体内过表达NLRP6可以显著降低血清中谷丙转氨酶和天冬氨酸转氨酶水平[34]。NLRP6可能通过抑制NF-κB信号通路降低人肝星状细胞LX-2细胞的活化、增殖和迁移能力,从而减少酒精性肝炎诱导的肝纤维化。在非酒精性脂肪性感染小鼠模型中,NLRP6缺失同样会加重肝脏脂肪变性、脂质代谢失调和纤维化,促进炎症发生[33]。以上研究表明,NLRP6可以防止肝脏疾病进一步发展,维持机体稳态。
3.3 NLRP6与中枢神经系统疾病炎症小体在中枢神经系统疾病中可引发无菌性炎症,如创伤性脑损伤(traumatic brain injury)、缺血性脑卒中(cerebral ischemic stroke)和脑内出血等疾病[59–61]。脑内出血是一种严重的神经系统疾病,小胶质细胞在脑内出血时,反应非常活跃,被认为是脑内出血后炎症反应的代表细胞[62]。氯化血红素处理小胶质细胞BV2后,NLRP6无论是mRNA水平还是在蛋白水平均高表达,随着氯化血红素浓度的增加,BV2细胞中,促炎细胞因子TNF-α和IL-6的表达水平也显著升高[28]。另有研究发现,NLRP6可能通过降低原发性高血压的遗传易感性来预防脑内出血的发生[63]。Meng等[64]利用大脑中动脉阻塞(middle cerebral artery occlusion/ reperfusion)模型模拟脑缺血再灌注损伤(cerebral ischemia-reperfusion injury)发现NLRP6沉默后,脑梗死体积和脑含水量均下降,神经功能得到恢复,脑缺血再灌注损伤引起的炎症级联反应受到抑制。该课题组进一步研究发现,在体外氧糖剥夺/复(OGD/R)星形胶质细胞模型中,敲除NLRP6可显著降低神经元凋亡率,提高神经元活力[65]。综上,NLRP6在中枢神经系统中可以感知损伤、介导炎症反应和参与相关疾病的调控。
4 小结NLRP6是近年来新发现的NLRs蛋白家族成员,且它是该蛋白家族中唯一被证实可以对机体产生负调控作用的蛋白,近年来成为研究的热点。对于不同病原体甚至在不同器官或细胞中,NLRP6表现出了截然相反的功能。机体是如何调控NLRP6参与炎症反应以及NLRP6在不同细胞和器官中所发挥的特异性功能仍需进一步探索。此外,已有研究表明趋化因子CXCL1和CXCL2能够通过G蛋白偶联受体CXCR2诱导NLRP3炎症小体激活[66]。缺失NLRP6的机体受到病原体感染时,这些趋化因子表达明显上调,推测NLRP6可能通过抑制趋化因子负调控NLRP3表达。总之,NLRP6在多种疾病发生发展中具有重要意义,对NLRP6的深入研究有利于为多种疾病提供新的治疗靶点和思路。
References
| [1] | Schaefer L. Complexity of danger: the diverse nature of damage-associated molecular patterns. Journal of Biological Chemistry, 2014, 289(51): 35237-35245. DOI:10.1074/jbc.R114.619304 |
| [2] | Tang DL, Kang R, Coyne CB, Zeh HJ, Lotze MT. PAMPs and DAMPs: signal 0s that spur autophagy and immunity. Immunological Reviews, 2012, 249(1): 158-175. DOI:10.1111/j.1600-065X.2012.01146.x |
| [3] | Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nature Immunology, 2010, 11(5): 373-384. DOI:10.1038/ni.1863 |
| [4] | Mukherjee S, Hooper LV. Antimicrobial defense of the intestine. Immunity, 2015, 42(1): 28-39. DOI:10.1016/j.immuni.2014.12.028 |
| [5] | Grenier JM, Wang L, Manji GA, Huang WJ, Al-Garawi A, Kelly R, Carlson A, Merriam S, Lora JM, Briskin M, DiStefano PS, Bertin J. Functional screening of five PYPAF family members identifies PYPAF5 as a novel regulator of NF-κB and caspase-1. FEBS Letters, 2002, 530(1/2/3): 73-78. |
| [6] | Kanneganti TD. Central roles of NLRs and inflammasomes in viral infection. Nature Reviews Immunology, 2010, 10(10): 688-698. DOI:10.1038/nri2851 |
| [7] | Lamkanfi M, Dixit VM. Mechanisms and functions of inflammasomes. Cell, 2014, 157(5): 1013-1022. DOI:10.1016/j.cell.2014.04.007 |
| [8] | Anand PK, Malireddi RKS, Lukens JR, Vogel P, Bertin J, Lamkanfi M, Kanneganti TD. NLRP6 negatively regulates innate immunity and host defence against bacterial pathogens. Nature, 2012, 488(7411): 389-393. DOI:10.1038/nature11250 |
| [9] | Chen GY, Liu MC, Wang FY, Bertin J, Núñez G. A functional role for Nlrp6 in intestinal inflammation and tumorigenesis. Journal of Immunology, 2011, 186(12): 7187-7194. DOI:10.4049/jimmunol.1100412 |
| [10] | Elinav E, Strowig T, Kau AL, Henao-Mejia J, Thaiss CA, Booth CJ, Peaper DR, Bertin J, Eisenbarth SC, Gordon JI, Flavell RA. NLRP6 inflammasome regulates colonic microbial ecology and risk for colitis. Cell, 2011, 145(5): 745-757. DOI:10.1016/j.cell.2011.04.022 |
| [11] | Normand S, Delanoye-Crespin A, Bressenot A, Huot L, Grandjean T, Peyrin-Biroulet L, Lemoine Y, Hot D, Chamaillard M. Nod-like receptor pyrin domain- containing protein 6 (NLRP6) controls epithelial self-renewal and colorectal carcinogenesis upon injury. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(23): 9601-9606. DOI:10.1073/pnas.1100981108 |
| [12] | Ydens E, Demon D, Lornet G, De Winter V, Timmerman V, Lamkanfi M, Janssens S. Nlrp6 promotes recovery after peripheral nerve injury independently of inflammasomes. Journal of Neuroinflammation, 2015, 12: 143. DOI:10.1186/s12974-015-0367-8 |
| [13] | Zheng DP, Kern L, Elinav E. The NLRP6 inflammasome. Immunology, 2021, 162(3): 281-289. DOI:10.1111/imm.13293 |
| [14] | Kayagaki N, Warming S, Lamkanfi M, Walle LV, Louie S, Dong J, Newton K, Qu Y, Liu JF, Heldens S, Zhang J, Lee WP, Roose-Girma M, Dixit VM. Non-canonical inflammasome activation targets caspase-11. Nature, 2011, 479(7371): 117-121. DOI:10.1038/nature10558 |
| [15] | Ding JJ, Wang K, Liu W, She Y, Sun Q, Shi JJ, Sun HZ, Wang DC, Shao F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature, 2016, 535(7610): 111-116. DOI:10.1038/nature18590 |
| [16] | Martinon F, Burns K, Tschopp J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Molecular Cell, 2002, 10(2): 417-426. DOI:10.1016/S1097-2765(02)00599-3 |
| [17] | Shen C, Lu A, Xie WJ, Ruan JB, Negro R, Egelman EH, Fu TM, Wu H. Molecular mechanism for NLRP6 inflammasome assembly and activation. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(6): 2052-2057. DOI:10.1073/pnas.1817221116 |
| [18] | Levy M, Thaiss CA, Zeevi D, Dohnalová L, Zilberman-Schapira G, Mahdi JA, David E, Savidor A, Korem T, Herzig Y, Pevsner-Fischer M, Shapiro H, Christ A, Harmelin A, Halpern Z, Latz E, Flavell RA, Amit I, Elinav E. Microbiota-modulated metabolites shape the intestinal microenvironment by regulating NLRP6 inflammasome signaling. Cell, 2015, 163(6): 1428-1443. DOI:10.1016/j.cell.2015.10.048 |
| [19] | Wang G, Wang QB, Bai J, Zhao N, Wang Y, Zhou R, Kong W, Zeng TS, Tao KX, Wang GB, Xia ZF. Upregulation of intestinal NLRP6 inflammasomes after roux-en-Y gastric bypass promotes gut immune homeostasis. Obesity Surgery, 2020, 30(1): 327-335. DOI:10.1007/s11695-019-04152-4 |
| [20] | Hara H, Seregin SS, Yang DH, Fukase K, Chamaillard M, Alnemri ES, Inohara N, Chen GY, Núñez G. The NLRP6 inflammasome recognizes lipoteichoic acid and regulates Gram-positive pathogen infection. Cell, 2018, 175(6): 1651-1664.e14. DOI:10.1016/j.cell.2018.09.047 |
| [21] | Leng FW, Yin H, Qin SY, Zhang K, Guan YK, Fang R, Wang HL, Li GH, Jiang ZF, Sun F, Wang DC, Xie C. NLRP6 self-assembles into a linear molecular platform following LPS binding and ATP stimulation. Scientific Reports, 2020, 10(1): 198. DOI:10.1038/s41598-019-57043-0 |
| [22] | Lu WL, Zhang L, Song DZ, Yi XW, Xu WZ, Ye L, Huang DM. NLRP6 suppresses the inflammatory response of human periodontal ligament cells by inhibiting NF-κB and ERK signal pathways. International Endodontic Journal, 2019, 52(7): 999-1009. DOI:10.1111/iej.13091 |
| [23] | Mukherjee S, Kumar R, Tsakem Lenou E, Basrur V, Kontoyiannis DL, Ioakeimidis F, Mosialos G, Theiss AL, Flavell RA, Venuprasad K. Deubiquitination of NLRP6 inflammasome by Cyld critically regulates intestinal inflammation. Nature Immunology, 2020, 21(6): 626-635. DOI:10.1038/s41590-020-0681-x |
| [24] | Kempster SL, Belteki G, Forhead AJ, Fowden AL, Catalano RD, Lam BY, McFarlane I, Charnock-Jones DS, Smith GCS. Developmental control of the Nlrp6 inflammasome and a substrate, IL-18, in mammalian intestine. American Journal of Physiology Gastrointestinal and Liver Physiology, 2011, 300(2): G253-G263. DOI:10.1152/ajpgi.00397.2010 |
| [25] | Sun YD, Zhang M, Chen CC, Gillilland M Ⅲ, Sun X, El-Zaatari M, Huffnagle GB, Young VB, Zhang, Hong SC, Chang YM, Gumucio DL, Owyang C, Kao JY. Stress-induced corticotropin-releasing hormone-mediated NLRP6 inflammasome inhibition and transmissible enteritis in mice. Gastroenterology, 2013, 144(7): 1478-1487.e8. DOI:10.1053/j.gastro.2013.02.038 |
| [26] | Mao XQ, Qiu XY, Jiao CH, Lu MJ, Zhao XJ, Li XT, Li JJ, Ma JJ, Zhang HJ. Candida albicans SC5314 inhibits NLRP3/NLRP6 inflammasome expression and dampens human intestinal barrier activity in Caco-2 cell monolayer model. Cytokine, 2020, 126: 154882. DOI:10.1016/j.cyto.2019.154882 |
| [27] | Xu XW, Zhu X, Wang CY, Li Y, Fan CG, Kao XM. microRNA-650 promotes inflammation induced apoptosis of intestinal epithelioid cells by targeting NLRP6. Biochemical and Biophysical Research Communications, 2019, 517(4): 551-556. DOI:10.1016/j.bbrc.2019.06.077 |
| [28] | Nie H, Hu Y, Guo WL, Wang WZ, Yang QW, Dong Q, Tang YP, Li Q, Tang ZP. miR-331-3p inhibits inflammatory response after intracerebral hemorrhage by directly targeting NLRP6. BioMed Research International, 2020, 2020: 6182464. |
| [29] | Bai YL, Li S. Long noncoding RNA OIP5-AS1 aggravates cell proliferation, migration in gastric cancer by epigenetically silencing NLRP6 expression via binding EZH2. Journal of Cellular Biochemistry, 2020, 121(1): 353-362. DOI:10.1002/jcb.29183 |
| [30] | Wang QQ, Wang CM, Chen JL. NLRP6, decreased in gastric cancer, suppresses tumorigenicity of gastric cancer cells. Cancer Management and Research, 2018, 10: 6431-6444. DOI:10.2147/CMAR.S182980 |
| [31] | Yu LM, Zhao KJ, Wang SS, Wang X, Lu B. Corticotropin-releasing factor induces inflammatory cytokines via the NLRP6-inflammatory cytokine axis in a murine model of irritable bowel syndrome. Journal of Digestive Diseases, 2019, 20(3): 143-151. DOI:10.1111/1751-2980.12704 |
| [32] | Anand PK, Kanneganti TD. NLRP6 in infection and inflammation. Microbes and Infection, 2013, 15(10/11): 661-668. |
| [33] | Huang CY, Liu QH, Tang Q, Jing XD, Wu T, Zhang JH, Zhang GR, Zhou J, Zhang ZJ, Zhao YN, Huang H, Xia Y, Yan JM, Xiao J, Li YP, He JH. Hepatocyte-specific deletion of Nlrp6 in mice exacerbates the development of non-alcoholic steatohepatitis. Free Radical Biology & Medicine, 2021, 169: 110-121. |
| [34] | Ji XY, Li LL, Lu PP, Li X, Tian DA, Liu M. NLRP6 exerts a protective role via NF-kB with involvement of CCL20 in a mouse model of alcoholic hepatitis. Biochemical and Biophysical Research Communications, 2020, 528(3): 485-492. DOI:10.1016/j.bbrc.2020.05.171 |
| [35] | Xu DY, Wu XP, Peng LC, Chen TT, Huang QY, Wang Y, Ye C, Peng YY, Hu DL, Fang RD. The critical role of NLRP6 inflammasome in Streptococcus pneumoniae infection in vitro and in vivo. International Journal of Molecular Sciences, 2021, 22(8): 3876. DOI:10.3390/ijms22083876 |
| [36] | Li RZ, Zhu S. NLRP6 inflammasome. Molecular Aspects of Medicine, 2020, 76: 100859. DOI:10.1016/j.mam.2020.100859 |
| [37] | Chen H, Li YH, Gu JJ, Yin L, Bian F, Su LS, Hong YH, Deng Y, Chi W. TLR4-MyD88 pathway promotes the imbalanced activation of NLRP3/NLRP6 via caspase-8 stimulation after alkali burn injury. Experimental Eye Research, 2018, 176: 59-68. DOI:10.1016/j.exer.2018.07.001 |
| [38] | Li MF, Chen YT, Shi JR, Ju W, Qi K, Fu CL, Li ZY, Zhang X, Qiao JL, Xu KL, Zeng LY. NLRP6 deficiency aggravates liver injury after allogeneic hematopoietic stem cell transplantation. International Immunopharmacology, 2019, 74: 105740. DOI:10.1016/j.intimp.2019.105740 |
| [39] | Girardin SE, Boneca IG, Carneiro LAM, Antignac A, Jéhanno M, Viala J, Tedin K, Taha MK, Labigne A, Zähringer U, Coyle AJ, DiStefano PS, Bertin J, Sansonetti PJ, Philpott DJ. Nod1 detects a unique muropeptide from Gram-negative bacterial peptidoglycan. Science, 2003, 300(5625): 1584-1587. DOI:10.1126/science.1084677 |
| [40] | Gross O, Poeck H, Bscheider M, Dostert C, Hannesschläger N, Endres S, Hartmann G, Tardivel A, Schweighoffer E, Tybulewicz V, Mocsai A, Tschopp J, Ruland J. Syk kinase signalling couples to the Nlrp3 inflammasome for anti-fungal host defence. Nature, 2009, 459(7245): 433-436. DOI:10.1038/nature07965 |
| [41] | Kobayashi KS, Chamaillard M, Ogura Y, Henegariu O, Inohara N, Nuñez G, Flavell RA. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science, 2005, 307(5710): 731-734. DOI:10.1126/science.1104911 |
| [42] | Paudel S, Ghimire L, Jin LL, Baral P, Cai SS, Jeyaseelan S. NLRC4 suppresses IL-17A-mediated neutrophil-dependent host defense through upregulation of IL-18 and induction of necroptosis during Gram-positive pneumonia. Mucosal Immunology, 2019, 12(1): 247-257. DOI:10.1038/s41385-018-0088-2 |
| [43] | Ghimire L, Paudel S, Jin LL, Baral P, Cai SS, Jeyaseelan S. NLRP6 negatively regulates pulmonary host defense in Gram-positive bacterial infection through modulating neutrophil recruitment and function. PLoS Pathogens, 2018, 14(9): e1007308. DOI:10.1371/journal.ppat.1007308 |
| [44] | Cai SS, Paudel S, Jin LL, Ghimire L, Taylor CM, Wakamatsu N, Bhattarai D, Jeyaseelan S. NLRP6 modulates neutrophil homeostasis in bacterial pneumonia-derived Sepsis. Mucosal Immunology, 2021, 14(3): 574-584. DOI:10.1038/s41385-020-00357-4 |
| [45] | Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, Weinrauch Y, Zychlinsky A. Neutrophil extracellular traps kill bacteria. Science: New York, N Y, 2004, 303(5663): 1532-1535. DOI:10.1126/science.1092385 |
| [46] | Wlodarska M, Thaiss CA, Nowarski R, Henao-Mejia J, Zhang JP, Brown EM, Frankel G, Levy M, Katz MN, Philbrick WM, Elinav E, Finlay BB, Flavell RA. NLRP6 inflammasome orchestrates the colonic host-microbial interface by regulating goblet cell mucus secretion. Cell, 2014, 156(5): 1045-1059. DOI:10.1016/j.cell.2014.01.026 |
| [47] | Birchenough GM, Nyström EE, Johansson ME, Hansson GC. A sentinel goblet cell guards the colonic crypt by triggering Nlrp6-dependent Muc2 secretion. Science: New York, N Y, 2016, 352(6293): 1535-1542. DOI:10.1126/science.aaf7419 |
| [48] | Wang PH, Zhu S, Yang L, Cui S, Pan W, Jackson R, Zheng YJ, Rongvaux A, Sun QM, Yang G, Gao SD, Lin RT, You FP, Flavell R, Fikrig E. Nlrp6 regulates intestinal antiviral innate immunity. Science, 2015, 350(6262): 826-830. DOI:10.1126/science.aab3145 |
| [49] | Zhu S, Ding SY, Wang PH, Wei Z, Pan W, Palm NW, Yang Y, Yu H, Li HB, Wang G, Lei XQ, de Zoete MR, Zhao J, Zheng YJ, Chen HW, Zhao YJ, Jurado KA, Feng NG, Shan L, Kluger Y, Lu J, Abraham C, Fikrig E, Greenberg HB, Flavell RA. Nlrp9b inflammasome restricts Rotavirus infection in intestinal epithelial cells. Nature, 2017, 546(7660): 667-670. DOI:10.1038/nature22967 |
| [50] | Sateriale A, Gullicksrud JA, Engiles JB, McLeod BI, Kugler EM, Henao-Mejia J, Zhou T, Ring AM, Brodsky IE, Hunter CA, Striepen B. The intestinal parasite Cryptosporidium is controlled by an enterocyte intrinsic inflammasome that depends on NLRP6. Proceedings of the National Academy of Sciences of the United States of America, 2021, 118(2): e2007807118. DOI:10.1073/pnas.2007807118 |
| [51] | Wang JF, Dong R, Zheng S. Roles of the inflammasome in the gut-liver axis (Review). Molecular Medicine Reports, 2019, 19(1): 3-14. |
| [52] | Levy M, Shapiro H, Thaiss CA, Elinav E. NLRP6: a multifaceted innate immune sensor. Trends in Immunology, 2017, 38(4): 248-260. DOI:10.1016/j.it.2017.01.001 |
| [53] | Seregin SS, Golovchenko N, Schaf B, Chen JC, Pudlo NA, Mitchell J, Baxter NT, Zhao LL, Schloss PD, Martens EC, Eaton KA, Chen GY. NLRP6 protects Il10−/− mice from colitis by limiting colonization of Akkermansia muciniphila. Cell Reports, 2017, 19(4): 733-745. DOI:10.1016/j.celrep.2017.03.080 |
| [54] |
Wen Z, Tang ZH. Research progress of NLRP6. Chinese Journal of Clinicians: Electronic Edition, 2014, 8(15): 2847-2851.
(in Chinese) 温哲, 唐朝晖. NLRP6的研究进展. 中华临床医师杂: 电子版, 2014, 8(15): 2847-2851. |
| [55] | Seregin SS, Golovchenko N, Schaf B, Chen J, Eaton KA, Chen GY. NLRP6 function in inflammatory monocytes reduces susceptibility to chemically induced intestinal injury. Mucosal Immunology, 2017, 10(2): 434-445. DOI:10.1038/mi.2016.55 |
| [56] | Tomuschat C, Virbel CR, O'Donnell AM, Puri P. Reduced expression of the NLRP6 inflammasome in the colon of patients with Hirschsprung's disease. Journal of Pediatric Surgery, 2019, 54(8): 1573-1577. DOI:10.1016/j.jpedsurg.2018.08.059 |
| [57] | Henao-Mejia J, Elinav E, Jin CC, Hao LM, Mehal WZ, Strowig T, Thaiss CA, Kau AL, Eisenbarth SC, Jurczak MJ, Camporez JP, Shulman GI, Gordon JI, Hoffman HM, Flavell RA. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature, 2012, 482(7384): 179-185. DOI:10.1038/nature10809 |
| [58] | Copelan EA. Hematopoietic stem-cell transplantation. New England Journal of Medicine, 2006, 354(17): 1813-1826. DOI:10.1056/NEJMra052638 |
| [59] | Abulafia DP, De Rivero Vaccari JP, Lozano JD, Lotocki G, Keane RW, Dietrich WD. Inhibition of the inflammasome complex reduces the inflammatory response after thromboembolic stroke in mice. Journal of Cerebral Blood Flow and Metabolism, 2009, 29(3): 534-544. DOI:10.1038/jcbfm.2008.143 |
| [60] | Ma QY, Chen S, Hu Q, Feng H, Zhang JH, Tang JP. NLRP3 inflammasome contributes to inflammation after intracerebral hemorrhage. Annals of Neurology, 2014, 75(2): 209-219. DOI:10.1002/ana.24070 |
| [61] | Tomura S, de Rivero Vaccari JP, Keane RW, Bramlett HM, Dietrich WD. Effects of therapeutic hypothermia on inflammasome signaling after traumatic brain injury. Journal of Cerebral Blood Flow & Metabolism, 2012, 32(10): 1939-1947. |
| [62] | Zhao XR, Sun GH, Zhang J, Strong R, Song WT, Gonzales N, Grotta JC, Aronowski J. Hematoma resolution as a target for intracerebral hemorrhage treatment: role for peroxisome proliferator-activated receptor γ in microglia/macrophages. Annals of Neurology, 2007, 61(4): 352-362. DOI:10.1002/ana.21097 |
| [63] | Herrera VLM, Bagamasbad P, Didishvili T, Decano JL, Ruiz-Opazo N. Overlapping genes in Nalp6/PYPAF5 locus encode two V2-type vasopressin isoreceptors: angiotensin-vasopressin receptor (AVR) and non-AVR. Physiological Genomics, 2008, 34(1): 65-77. DOI:10.1152/physiolgenomics.00199.2007 |
| [64] | Meng CC, Zhang JY, Zhang LY, Wang YT, Li ZY, Zhao J. Effects of NLRP6 in cerebral ischemia/reperfusion (I/R) injury in rats. Journal of Molecular Neuroscience, 2019, 69(3): 411-418. DOI:10.1007/s12031-019-01370-4 |
| [65] | Zhang JY, Jiang N, Zhang LY, Meng CC, Zhao J, Wu JX. NLRP6 expressed in astrocytes aggravates neurons injury after OGD/R through activating the inflammasome and inducing pyroptosis. International Immunopharmacology, 2020, 80: 106183. DOI:10.1016/j.intimp.2019.106183 |
| [66] | Boro M, Balaji KN. CXCL1 and CXCL2 regulate NLRP3 inflammasome activation via G-protein-coupled receptor CXCR2. Journal of Immunology, 2017, 199(5): 1660-1671. DOI:10.4049/jimmunol.1700129 |