 2022, Vol. 62
2022, Vol. 62
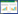


海底热液环境中嗜中性微需氧铁氧化菌的多样性、生物矿化作用及其代谢特征

http://dx.doi.org/10.13343/j.cnki.wsxb.20210639
中国科学院微生物研究所,中国微生物学会
中国科学院微生物研究所,中国微生物学会
文章信息
- 孙明雪, 宿蕾, 李江涛. 2022
- SUN Mingxue, SU Lei, LI Jiangtao.
- 海底热液环境中嗜中性微需氧铁氧化菌的多样性、生物矿化作用及其代谢特征
- Diversity, biomineralization, and metabolic characteristics of neutrophilic microaerophilic iron-oxidizing bacteria in seafloor hydrothermal environments
- 微生物学报, 62(6): 2119-2135
- Acta Microbiologica Sinica, 62(6): 2119-2135
-
文章历史
- 收稿日期:2021-10-22
- 修回日期:2021-12-21
- 网络出版日期:2022-03-12
引用本文 |
孙明雪, 宿蕾, 李江涛. 海底热液环境中嗜中性微需氧铁氧化菌的多样性、生物矿化作用及其代谢特征[J]. 微生物学报, 2022, 62(6): 2119-2135.
Sun Mingxue, Su Lei, Li Jiangtao. Diversity, biomineralization, and metabolic characteristics of neutrophilic microaerophilic iron-oxidizing bacteria in seafloor hydrothermal environments[J]. Acta Microbiologica Sinica, 2022, 62(6): 2119-2135.
海底热液环境中嗜中性微需氧铁氧化菌的多样性、生物矿化作用及其代谢特征
1. 海洋地质国家重点实验室(同济大学), 上海 200092;
2. 同济大学海洋与地球科学学院, 上海 200092
2. 同济大学海洋与地球科学学院, 上海 200092
摘要:铁元素是深海热液活动产物的主要成分之一,也是热液喷口处化能自养微生物生态系统的重要驱动元素。以Zetaproteobacteria为典型代表的嗜中性微需氧铁氧化菌是海底喷口及其周围环境中生物介导的Fe2+氧化这一生物矿化作用的主要驱动者。这些铁氧化菌通过氧化Fe2+获取维持自身代谢所必需的能量,同时分泌有机质将氧化后的不溶铁(氧化物或氢氧化物)沉淀于细胞外,形成具有螺旋丝带状、中空长杆状、分叉管状以及其他具有特殊形貌特征的显微结构体,进而堆积成广泛分布于海底的富铁氧化物/氢氧化物。越来越多的研究表明,编码细胞色素孔蛋白的cyc2基因是Zetaproteobacteria铁氧化菌进行Fe2+氧化的关键基因,而细胞色素c或其他周质细胞色素则是Fe2+氧化过程中的关键电子传递载体。基于宏基因组分析的系列研究揭示了Zetaproteobacteria普遍具有多种与氮、硫、氢以及砷元素循环密切相关的功能基因与代谢途径,暗示了其在上述元素循环过程中的潜在作用。本文系统地总结了海底热液喷口及其周围环境中发现的嗜中性微需氧铁氧化菌的多样性和生理特征以及它们进行Fe2+氧化生物矿化作用而形成的显微结构记录、介导Fe2+氧化的关键基因与电子传递路径等,有利于全面、系统地认识该类微生物在海底热液喷口关键成矿元素的迁移与富集、维系喷口生态以及微生物成矿等方面所起的作用与贡献。
关键词:海底热液系统 嗜中性微需氧铁氧化菌 Zetaproteobacteria纲 富铁显微结构 铁氧化机制 代谢特征
Diversity, biomineralization, and metabolic characteristics of neutrophilic microaerophilic iron-oxidizing bacteria in seafloor hydrothermal environments
1. State Key Laboratory of Marine Geology, Tongji University, Shanghai 200092, China;
2. School of Ocean and Earth Science, Tongji University, Shanghai 200092, China
2. School of Ocean and Earth Science, Tongji University, Shanghai 200092, China
Abstract: Iron is one of the main components of deep-sea hydrothermal activity products and an important driving element for the chemoautotrophic microbial ecosystems at hydrothermal vents. The neutrophilic microaerophilic iron-oxidizing bacteria, represented by Zetaproteobacteria, are the main drivers of biomediated Fe2+ oxidation in hydrothermal vents and their surroundings. Iron-oxidizing bacteria acquires the energy essential to maintain their metabolism through Fe2+ oxidation, while secrete organic matter to precipitate the oxidized insoluble iron (oxides or hydroxides) outside the cells, forming microstructures with twisted stalks, hollow sheaths, branching hollow tubes or other special morphological features. These microstructures accumulate into iron-rich oxides/hydroxides widely distributed on the seafloor. Increasing studies have demonstrated that cyc2, encoding cytochrome-porin, is the key gene of Fe2+ oxidation by Zetaproteobacteria, while c-type cytochromes or other periplasmic cytochromes are the main electron transport carriers in Fe2+ oxidation. The metagenome-based studies reveal that Zetaproteobacteria generally possess multiple functional genes and metabolic pathways associated with nitrogen, sulfur, hydrogen, and arsenic cycling, suggesting the potential role of Zetaproteobacteria in the cycling of the above elements. In this paper, we systematically summarized the diversity, physiological characteristics, biomineralization-formed microstructure records, and key genes and electron transport pathways that mediated Fe2+ oxidation of neutrophilic microaerophilic iron-oxidizing bacteria in hydrothermal environments. This review facilitates the systematical understanding about the role of these microorganisms in the migration and enrichment of key ore-forming elements, the maintenance of ecological balance, and the mineralization by microorganisms in submarine hydrothermal vents.
Keywords:
submarine hydrothermal systems neutrophilic microaerophilic iron-oxidizing bacteria Zetaproteobacteria iron-rich microstructure iron oxidation mechanism metabolic characteristics
铁元素是地球上最丰富的元素之一[1],在自然环境中主要以+2、+3价两种氧化还原价态存在,可以与碳、氧、氮、硫等元素形成复合物,广泛存在于多种矿物相中,也是生命活动必不可少的关键元素[2–3]。Fe2+在空气中极易发生化学氧化,但在O2浓度范围为5–50 μmol/L的微氧环境中却可以被一些嗜中性(pH 6.0–7.5)的铁氧化菌(Fe-oxidizing bacteria,FeOB)利用进行化能自养生活[4–7]。典型的嗜中性微需氧FeOB在19世纪被首次发现[8],它们通常生活在有氧–无氧过渡界面的富铁含水环境中,包括湿地、地下水、海洋沉积物以及海底热液喷口等[5, 9]。大多数嗜中性微需氧的FeOB会产生具有特殊形貌特征的、主要由有机物和铁氧化物/氢氧化物组成的螺旋丝带状或中空长杆状显微结构体[5, 9–11]。从系统发育分类上来说,目前发现的这些铁氧化微生物主要隶属于Proteobacteria门,包括栖息于淡水环境的Gammaproteobacteria和海洋环境中发现的Zetaproteobacteria。其中,前者主要包括Gallionella、Leptothrix、Sideroxydans、Ferriphaselus和Ferritrophicum等,后者则主要由Mariprofundus和Ghiorsea构成[5, 12–14]。目前为止,所有隶属于Zetaproteobacteria的纯培养微生物都能从Fe2+氧化这一过程中获取能量,而基于基因组的分析也表明所有的Zetaproteobacteria类群都具有微需氧条件下进行Fe2+氧化的代谢潜能[13–14]。Fe2+是海底热液的主要化学成分之一,Fe2+氧化也是热液喷口及其周围环境中嗜中性微需氧铁氧化菌的主要能量来源[14]。特别是在低温弥散流区域(t < 100 ℃),由铁氧化菌介导的Fe2+氧化这一生物矿化作用是低温弥散流喷溢过程中形成的富铁氧化物/氢氧化物的最主要的成因。这些富铁氧化物/氢氧化物已被发现广泛分布于全球的海底,包括海山、洋中脊、弧后扩张中心、板内火山活动以及大陆边缘断裂带等多种不同的构造环境[15–17],也意味着嗜中性微需氧铁氧化菌在全球海底普遍分布。在过去的近20年里,人们已经尝试对海底热液喷口环境中的铁氧化微生物及其Fe2+氧化的生物矿化作用展开系统的研究,结合铁氧化菌的分离、培养与组学分析等,在铁氧化菌的多样性、生理特性以及生物矿化机制方面取得了重要的进展。本文对上述研究进展进行了系统的总结,以期能够加深人们对海底热液喷口及其周围环境中栖息的嗜中性微需氧铁氧化菌的认识。
1 嗜中性微需氧铁氧化菌的多样性早在19世纪中期,人们就发现在陆地富含Fe2+的淡水中存在着铁氧化菌,它们隶属于Gallionella和Leptothrix两个属,通常会形成典型的螺旋丝带状和中空长杆状等可以用来识别这些铁氧化菌的标志性富铁结构[5, 18–19]。20世纪80年代,科学家们首次从海洋中发现了上述螺旋状和长杆状结构,并据此推断这些结构是由Gallionella spp.和Leptothrix spp. 形成的,因而进一步推测这两类铁氧化菌也可以生活在海洋中[20]。然而,直到2002年,基于原核生物SSU rRNA基因序列的系统发育分析表明这类特征性的富铁显微结构并不是由隶属于Gallionella和Leptothrix的铁氧化菌形成的,而是来自于一类以Mariprofundus ferrooxydans为代表的新的FeOB矿化产物,后来将这些FeOB命名为一个新的类群,即Zetaproteobacteria[15, 20]。
Zetaproteobacteria是在海底热液喷口及其周围环境中最常见、也是分布最广的铁氧化微生物类群[5, 13]。Zetaproteobacteria属于典型的嗜中性、微需氧、化能自养型FeOB,最初是从夏威夷Lō’ihi海山的低温热液喷口附近发现、分离并被命名的[15, 20]。在氧气浓度充足且pH近中性的情况下,Fe2+的无机化学氧化速率非常快,微生物难以与之竞争,只有在微氧环境(O2 < 50 µmol/L)中,FeOB氧化Fe2+的速率才能够超过非生物氧化的速率。因此,Zetaproteobacteria铁氧化菌大多数都分布于深海或近岸环境的有氧—无氧界面附近[4, 7]。除了Lō’ihi海山,几乎全球海底的热液喷口周围的微生物菌席(图 1A–C)、低温热液沉淀物或氧化物(图 1D)、热液喷口宏生物的共生菌以及喷口周围的玄武岩中都发现了Zetaproteobacteria[21–27]。此外,在沿海近岸的海洋沉积物、地下水和氧化还原分层水柱以及陆地上富含CO2的深层地下水、沼泽湿地等富铁环境中也陆续发现了Zetaproteobacteria铁氧化菌存在的证据,表明它们的分布范围广泛,并不局限于深海热液喷口环境[9, 28–33]。在上述Zetaproteobacteria分布的环境中,Fe2+浓度通常可以达到数百µmol/L,然而,近些年来,随着研究范围的增加,在沿海一些低Fe2+浓度(几十µmol/L甚至低于0.2 µmol/L)的环境中,也发现了Zetaproteobacteria铁氧化菌的分布[9]。

|
| 图 1 海底热液喷口嗜中性微需氧铁氧化菌的常见栖息形式 Figure 1 Common habitats of neutrophilic, microaerophilic Fe(Ⅱ)-oxidizing bacteria in seafloor hydrothermal vents. A: iron microbial mats on the surface of sulfide chimney (yellow)[13]; B: curd-type mats formed by FeOB around the hydrothermal vents[13]; C: veil-type mats formed by FeOB around the vents[13]; D: iron-silica-rich low-temperature hydrothermal precipitates formed by FeOB. A from the Urashima vent site, Mariana Trough[13]; B–C from Lō'ihi seamount in Hawaii[13]; D from the Longqi hydrothermal field, the Southwest Indian ridge, photographed by authors' group. |
一直以来,从深海环境中获得FeOB的纯培养是开展相关研究所面临的主要挑战之一[13, 34]。琼脂凝胶反向梯度管培养法是当前分离微需氧FeOB最常用的方法,即在培养管中提供具有相反浓度梯度的Fe2+和O2浓度条件,让FeOB自行寻找最适生长位置[35]。目前,人们已从海洋环境中分离得到了21株纯培养的Zetaproteobacteria铁氧化菌(表 1和图 2),它们分别被划归为Mariprofundas和Ghiorsea属,在这21株纯菌中,有7株是从海底热液相关的环境中分离得到的[13–14, 20, 28, 34, 36–41]。纯培养菌株Mariprofundus ferrooxydans PV-1、JV-1、M34和Mariprofundus sp. EKF-M39均分离自夏威夷Lō’ihi海山的低温弥散流热液区,均为专性铁氧化细菌,革兰氏阴性菌,细胞为豆状,且都能够形成富铁的螺旋丝带状结构体(如图 3A和3H)[11, 20, 34, 36, 42]。Mariprofundus micogutta ET2分离自伊豆—小笠原岛弧Bayonnaise海丘附近的热液沉积物,也是专性铁氧化微生物,它的细胞通常是长度为1–2 μm的豆状或弯曲杆状,会在胞外形成细丝状的富铁显微结构(图 3E)[39]。Ghiorsea bivora TAG-1与G. bivora SV-108分别是从大西洋洋中脊和马里亚纳海沟的热液喷口分离得到的。与Mariprofundus spp.不同,这2株菌属于兼性铁氧化细菌,除了可以进行Fe2+氧化之外,还可以利用H2氧化进行生长,其代谢活动形成的富铁显微结构通常为无定形状(图 3J)[14]。
表 1. 已获得分离培养的Zetaproteobacteria铁氧化菌(据参考文献[13]修改、补充)
Table 1. Isolates of the Zetaproteobacteria (modified and supplemented after reference [13])
| Isolate | Isolation source | ZetaOTU | Primary biomineral morphology | Sole electron donor | References |
| Mariprofundus ferrooxydans PV-1 | Lō’ihi hydrothermal vents | ZetaOTU11 | Stalk | Fe2+ | [20] |
| Mariprofundus ferrooxydans JV-1 | Lō’ihi hydrothermal vents | ZetaOTU11 | Stalk | Fe2+ | [20] |
| Mariprofundus ferrooxydans M34 | Lō’ihi hydrothermal vents | ZetaOTU11 | Stalk | Fe2+ | [34] |
| Mariprofundus sp. EKF-M39 | Lō’ihi hydrothermal vents | ZetaOTU36 | Stalk | Fe2+ | [36] |
| Mariprofundus micogutta ET2 | Bayonnaise hydrothermal | ZetaOTU18 | Thin filaments | Fe2+ | [39] |
| Ghiorsea bivora TAG-1 | Mid-Atlantic ridge hydrothermal vents | ZetaOTU9 | None | Fe2+ or H2 | [14] |
| Ghiorsea bivora SV-108 | Mariana hydrothermal vents | ZetaOTU9 | None | Fe2+ or H2 | [14] |
| Mariprofundus ferrinatatus CP-5 | Chesapeake Bay stratified water column | ZetaOTU18 | Dreads only | Fe2+ | [29] |
| Mariprofundus aestuarium CP-8 | Chesapeake Bay stratified water column | ZetaOTU37 | Dreads only | Fe2+ | [29] |
| Mariprofundus sp. GSB-2 | Great Salt Bay salt marsh Fe mat | ZetaOTU23 | Stalk | Fe2+ | [54] |
| Zetaproteobacteria sp. NF | Norsminde Fjord estuary sediments | ZetaOTU3 | Stalk | Fe2+ | [9] |
| Zetaproteobacteria sp. KV | Kalø Vig beach sediments | ZetaOTU3 | Stalk | Fe2+ | [9] |
| Zetaproteobacteria sp. CSS-1 | Coastal sediment bloodworm microcosm | ZetaOTU14 | Stalk | Fe2+ | [28] |
| Zetaproteobacteria sp. SR-1* | Worm burrow in Sheepscot estuary sediments | ZetaOTU3 | Stalk | Fe2+ | [28] |
| Zetaproteobacteria sp. ECHO-1* | Worm burrow in Sheepscot estuary sediments | ZetaOTU55 | None | Fe2+ | [28] |
| Mariprofundus ferrooxydans SC-2 | Big Fisherman’s Cove pyrrhotite colonization | ZetaOTU11 | Stalk | Fe2+ | [55] |
| Mariprofundus sp. EBB-1 | Damariscotta estuary pyrrhotite colonization | ZetaOTU14 | Stalk | Fe2+ | [40] |
| Mariprofundus sp. DIS-1 | West Boothbay Harbor mild steel incubation | ZetaOTU18 | Stalk | Fe2+ | [56] |
| Mariprofundus erugo P3 | Mallard Creek 316 stainless steel incubation | New ZetaOTU | Stalk | Fe2+ | [37] |
| Mariprofundus erugo P7 | North Creek 316 stainless steel incubation | New ZetaOTU | Stalk | Fe2+ | [37] |
| Mariprofundus ferrooxydans O-1 | Pamlico Sound steel-hulled shipwreck | ZetaOTU11 | Stalk | Fe2+ | [41] |
| *: reference from [28], but there was no named description in this paper, and the named description information reference from [13, 57]. | |||||

|
| 图 2 已经获得纯培养的Zetaproteobacteria铁氧化菌的系统发育树 Figure 2 Phylogenetic tree of Zetaproteobacteria isolates. The tree constructed by MEGA X maximum likelihood method[53]; bootstrap values were calculated from 1 000 replications, and only > 50% were reported; the codes in the parentheses represent the sequence record numbers in the common database; 16S rRNA gene sequences were obtained from NCBI and JGI databases. |

|
| 图 3 海底热液喷口及其周围环境中常见的由Zetaproteobacteria形成的特征性显微结构 Figure 3 Common characteristic microscopic structures formed by Zetaproteobacteria in seafloor hydrothermal vents and their surroundings. A: twisted stalks with directional arrangement[10]; B: semi-parallel directional long hollow sheaths; C: short branching hollow tubes; D: nest-type biominerals attached to long sheaths[10]; E: filaments structures[39, 65]; F: short dreads structures (inferred cell position indicated by the yellow oval)[29]; G: the fibrils on the surface of long hollow sheaths[13]; H: transmission electron microscopy image of twisted stalk, which are composed of multiple parallel fibrils, and the white arrow indicating the cell of FeOB[11]; I: Y-shaped hollow tube, made up of multiple parallel fibrils, and the white arrow points to cells[10]; J: amorphous oxides formed by FeOB, shown in green as FeOB cells[14]. Images A, D, G, I from Lō'ihi seamount in Hawaii; B–C from Lau Basin, photographed by our research group[59]; E–F, H–I were the microstructure images of isolated strains M. micogutta ET2, M. aestuarium CP-8, M. ferrooxydans PV-1 and G. bivora TAG-1, respectively. |
随着研究的逐渐深入和不断积累,近些年来,研究人员对全球范围内获取的Zetaproteobacteria铁氧化菌的16S rRNA基因序列进行了系统发育分析。基于97%的序列相似性将Zetaproteobacteria铁氧化菌的16S rRNA基因序列划分为60个操作分类单元(ZetaOTU1– OTU60),这些分类单元又被进一步划分为两个科,分别为Mariprofundaceae和暂未定名的family 2[13, 43]。研究发现,部分ZetaOTUs所代表的铁氧化菌广泛地分布于全球海底的富铁氧化物/氢氧化物中,如ZetaOTU1–4普遍存在于不同区域的大多数样品中[13, 34, 44]。相比之下,有一些ZetaOTUs所代表的铁氧化菌则展现出一定程度的环境偏好性。例如,近期McAllister等在其得到的27个铁氧化菌的ZetaOTUs中发现有17个ZetaOTUs表现出对喷口菌席生境的偏好性,而ZetaOTU6代表的铁氧化菌则偏好栖息在流动性强、温度较高的热液喷口附近[13, 44]。Duchinski等从马里亚纳火山弧热液区和夏威夷Lō’ihi海山热液区采集了29个样品进行分析,结果发现在马里亚纳热液区中检测到的ZetaOTUs组成与Lō’ihi海山具有显著差异,有相当一部分的ZetaOTUs表现出了一定的生物地理分区的物理隔离性,分别仅存在于Lō’ihi海山或马里亚纳热液区[44]。
除Zetaproteobacteria外,Edwards等从Juan de Fuca热液区分离到几株隶属于Gammaproteobacteria和Alphaproteobacteria的嗜中性化能自养铁氧化细菌,从系统发育关系表明,它们与Marinobacter aquaeoolei和Hyphomonas jannaschiana最为密切,可以在微需氧条件下或在硝酸盐存在的厌氧条件下生长,也会产生一些螺旋丝带状、分叉树枝状等典型铁氧化菌形成的结构[45]。Sudek等从Vailulu’u海山的菌席中也分离出多株属于Gammaproteobacteria和少量属于Alphaproteobacteria的异养铁氧化菌,它们主要在pH 7.5的条件下生长,少量菌株微需氧,系统发育分析发现它们与Pseudoalteromonas、Pseudomonas和Thioclava等物种紧密相关[46]。
陆地淡水环境中的微需氧嗜中性铁氧化菌Gallionella、Leptothrix和Sideroxydans也在海底低温热液喷口或者不活动的热液硫化物风化形成的铁氧化物中被检测到[25, 26, 47–51]。尽管含量不高,却表明上述微生物类群的分布范围不局限于陆地淡水环境。不过,需要指出的是,上述3种铁氧化菌的发现均是基于不可培养的微生物遗传物质的检测,至今没有从海底热液喷口及其周围环境中分离得到相关的纯培养。此外,在最近一项研究中,McAllister等从夏威夷Lō’ihi海山的低温热液微生物菌席中基于宏基因组的分箱技术获得了一个新的未培养铁氧化菌的基因组,命名为Candidatus Ferristratum sp.,分类上属于DTB120门。基因组分析表明,C. Ferristratum sp.属于厌氧铁氧化菌,依赖于硝酸盐呼吸来进行Fe2+氧化[52]。然而,该铁氧化菌仅具有NO3–还原的部分功能基因,并不能独立完成完整的反硝化过程。进一步基于转录组的研究发现,C. Ferristratum sp.是通过与Zetaproteobacteria铁氧化菌共生/共营养的方式来共同完成反硝化和铁氧化过程[52]。
2 嗜中性微需氧铁氧化菌形成的特征性显微结构如前文所述,铁氧化细菌通过氧化Fe2+的过程来获取维持自身代谢所必需的能量,该过程会在细胞外形成如螺旋丝带状、中空长杆状、分叉树枝状、丝缕状等具有非常特殊的形貌特征的富铁显微结构,这些结构成为了识别嗜中性微需氧FeOB的重要指示标志。矿物分析表明,这些特征性显微结构主要是由纳米尺度的、结晶程度较弱的水铁矿组成,后期随着时间的增长以及成岩作用的影响,会向结晶程度更高、更稳定的纤铁矿、针铁矿或赤铁矿转化[9–11, 58–59]。这些显微结构与FeOB的细胞以及其他微生物聚集在一起,形成蓬松多孔、厚度从毫米到几米不等的微生物菌席或者富铁氧化物/氢氧化物(图 1),广泛分布于海底热液喷口及其周围的环境中[5, 10, 60–61]。
2.1 螺旋丝带状结构螺旋丝带状结构是嗜中性铁氧化菌产生的最常见的标志性的显微结构(图 3A),分离自Lō’ihi海山菌席的M. ferrooxydans PV-1是产生该显微结构的典型代表菌株。PV-1的细胞为豆状或弯曲状,螺旋状丝带结构体产生于细胞凹面(图 3H),丝带结构远离细胞的一端固着于岩石或菌席等环境表面,细胞向高氧浓度方向移动,带动丝带结构向该方向生长[10–11, 62]。通常情况下,这些螺旋丝带状结构体由铁氧化物/氢氧化物和多糖类有机质组成,直径约为0.3–1.8 μm,由2–30个纳米级的原纤维平行排列构成,矿物晶体在其表面成核并向外生长,使丝带结构逐渐变厚,Fe/C比也逐渐增加[10–11]。通过产生丝带状结构体的方式,细胞能够前往微生境中Fe2+和O2浓度梯度最适宜的位置生长。丝带状结构会因为细胞的分裂产生两两分支,呈辐射状,随着丝带逐渐变厚,细胞会主动与之脱离,并在新的地方固定下来,继续产生新的丝带状结构[10, 62]。产生这种显微结构的FeOB能够在Fe2+和O2浓度梯度变化剧烈的生境中生存下来,它们喜欢栖息于更靠近热液喷口的微生物菌席中,这类菌席呈现凝乳状(curd-type,图 1B),螺旋状丝带结构在其中表现出很强的方向性(图 3A)[10]。
2.2 中空长杆状结构中空的长杆状结构是海底热液环境中铁氧化菌形成的另一种常见的显微结构(图 3B和3G)。虽然迄今为止尚没有从海洋环境中分离得到能够形成该显微结构的纯培养,但Chan等和Fleming等通过荧光原位杂交的方法确定了Zetaproteobacteria是形成这种显微结构的主要微生物类群[10, 63]。通常情况下,这些中空的长杆状结构的直径约为0.5–1.0 μm,长度从十几μm到数百μm不等。无论从形态、空间尺寸还是到物质组成,由Zetaproteobacteria铁氧化细菌形成的这些长杆状结构都与由陆地淡水环境中Leptothrix spp.铁氧化菌产生的杆状结构十分相似[10, 63]。然而,目前仅从部分海洋环境中发现极少量的Leptothrix类群,所以通常认为海洋环境中发现的这类中空的长杆状显微结构是Zetaproteobacteria铁氧化菌的产物。这类结构的形成通常是由多个细胞共同产生,杆状结构体的一端固着在生长环境的基质上,多个细胞在中空腔体中生长分裂,同时向非固着端移动,逐渐将长杆状结构体遗留在细胞的后面[10, 13]。中空长杆状结构多出现在面纱状菌席(veil-type,图 1C)中,这类菌席通常覆盖在喷口周围的硫化物、岩石或者老的菌席等离热液喷口相对较远、环境更稳定的位置(图 3B和3D),菌席内部Fe2+和O2浓度变化相对平缓[10]。长杆状结构在菌席的一定区域内呈平行或半平行生长,这可能与菌席内Fe2+和O2的分布或者细胞对菌席外部刺激的响应有关[10, 39, 63–64]。
2.3 分叉空心管状与巢状结构分叉空心管状结构也是Zetaproteobacteria铁氧化菌形成的比较常见的富铁显微结构之一(图 3C和3I)[16]。该结构宽约0.5–1.0 μm,长度相对较短,大约5–50 μm,呈扁平状,可以连续分叉,由多个铁氧化物/氢氧化物的原纤维组成[10]。与前文提到的螺旋丝带状结构类似,铁氧化菌的细胞生长于管状结构的一端,当细胞发生分裂的时候,与细胞相连接的管状结构也发生分叉,形成“Y”字型结构(图 3I)[10, 63]。
此外,一种类似球形的巢状结构也经常在热液喷口及其周围的铁氧化物中被观察到(图 3D),它们通常由细胞分泌的富含铁氧化物的原纤维细丝缠绕构成,铁氧化菌的细胞可能隐匿于巢状结构的中心,不过目前尚未能够确定形成该结构的微生物种群[5, 10],但通常也被认为是Zetaproteobacteria铁氧化菌生物矿化作用的产物。分叉空心管状与巢状结构通常分布于螺旋丝带状或者中空长杆状结构围成的孔隙中(图 3C–D)。这种特定的生态位选择可能是一种生存策略,也可能是因为形成它们的铁氧化菌更能适应菌席深处低O2浓度的环境[10]。
近些年来,新的FeOB类群不断地被分离培养出来,由它们形成的新的、具有非常特殊的形貌特征的富铁显微结构也被不断地发现,例如由M. micogutta ET2形成的细丝状结构(图 3E)、M. ferrinatatus CP-5与M. aestuarium CP-8形成的脏辫状结构(图 3F)以及G. bivora TAG-1和SV-108等铁氧化菌形成的无定形铁氧化物(图 3J)等[14, 29, 39]。在海底热液喷口及其周围富铁的环境中,上述各种具有特殊形貌、富铁的显微结构已成为指示嗜中性微需氧铁氧化菌存在/活动的重要证据,并且研究发现这些结构表面具有不同的表面纹理和超微结构,反映出生物矿化过程的更多细节同时可能指示了不同的地球化学/物理生态位[59]。
3 嗜中性微需氧铁氧化菌的生物矿化机制与代谢特征由于缺乏同位素以及可广泛应用的Fe2+氧化的遗传标记,近中性微氧条件下微生物介导的Fe2+氧化的过程往往难以追踪,加之FeOB纯培养的分离有一定的技术难度,人们对FeOB介导的Fe2+氧化这一微生物矿化的分子机制以及FeOB的代谢特征等问题仍然不清楚[13, 52]。但近些年来,随着获取的Zetaproteobacteria纯培养菌株的逐渐增加以及测序技术的快速发展,基于FeOB基因组、转录组和蛋白组的多组学分析使得人们对Zetaproteobacteria铁氧化菌的生理生化特性,特别是它们进行Fe2+氧化的分子机制、电子传递途径以及代谢潜能等,均取得了十分重要的进展[36, 57, 64, 66]。
3.1 Fe2+氧化的矿化机制与电子传递途径在铁氧化菌形成的显微结构部分提到,这些独特的结构通常由铁氧化物/氢氧化物和多糖类有机质组成。研究表明,Zetaproteobacteria铁氧化菌通过Fe2+氧化将电子向胞内传递,同时分泌有机质与氧化得到的Fe3+结合形成原纤维构成特征显微结构沉淀于胞外,并且为避免Fe3+在细胞内沉淀成矿从而损伤细胞,Fe2+氧化过程很可能发生在细胞膜外[10]。FeOB通过形成原纤维将不溶或者低溶解度的铁氧化物/氢氧化物聚集起来,矿物晶体在其表面成核并生长,从而避免细胞陷入矿物的围困[10, 65]。目前,我们对FeOB形成的特殊显微结构的化学成分、纳米级形态以及形成过程等方面有了更全面的认识,但形成这些结构潜在的遗传机制一直未得到解决。最新一项研究对16个Zetaproteobacteria分离菌株的泛基因组基于性状分析发现,存在一个候选基因簇与螺旋丝带状结构中有机基质的合成与分泌相关,它在形成这种结构的Zetaproteobacteria中高度保守[38]。基因簇包含6个基因sfz1–sfz6 (sfz: stalk formation in Zetaproteobacteria),推测sfz1、sfz2基因与胞外多糖合成及分泌相关,sfz4、sfz6可能与细胞壁合成和胞外多糖的水解有关,不过这些都是根据泛基因组数据结果推测得到的,需要实验进一步验证[38]。
在过去10多年的研究中,越来越多的证据表明细胞色素孔蛋白Cyc2是铁氧化菌执行Fe2+氧化的关键酶,编码该蛋白的cyc2基因目前被认为是最可能成为识别嗜中性铁氧化菌的标志基因,它几乎出现在所有已知Zetaproteobacteria的基因组中[57, 67]。Cyc2作为Fe2+氧化酶最早是在嗜酸性氧化亚铁硫杆菌这一类铁氧化菌中得到证实的,后来在嗜中性铁氧化菌Gallionella spp.中发现了Cyc2的弱同源物[68–69]。近年来,在海洋嗜中性铁氧化菌M. ferrooxydans PV-1的蛋白质组中也发现了丰富的Cyc2的同源物Cyc2 PV-1蛋白。基于宏基因组学和宏转录组学的研究进一步证实了cyc2基因在深海热液嗜中性铁氧化菌的菌席中高度表达,并且发现cyc2基因的表达会响应Fe2+浓度的变化而受到调节[28, 57, 66, 68–69]。Keffer等对Cyc2结构进行了分析预测并开展了Cyc2 PV-1蛋白的异源表达实验,结果证明了Cyc2是一种具有铁氧化功能的外膜蛋白,从而为嗜中性铁氧化菌Fe2+氧化过程发生在细胞膜外以及Cyc2是该过程的功能蛋白提供了确凿的生物化学证据[67]。
除了cyc2基因外,编码血红素细胞色素c的cyc1基因也被发现广泛分布于Zetaproteobacteria铁氧化菌的基因组中。Cyc1被认为是Fe2+氧化途径中进行电子传递的周质电子载体,不过研究也发现存在其他多种潜在的周质细胞色素电子载体[43, 57, 66]。细胞外膜Cyc2蛋白通过氧化Fe2+获得的电子,一方面通过Cyc1或其他周质电子载体传递到细胞内膜的末端氧化酶,并还原O2产生质子动力用于ATP的生成。目前,已经发现了3种类型的末端氧化酶,包括cbb3型细胞色素c氧化酶(分为近端和远端)、aa3型细胞色素c氧化酶和细胞色素bd-I泛醇氧化酶[43, 57, 66]。另一方面,电子也可以通过上述电子载体反向传输到细胞内膜上的bc1复合物或替代复合物ACIII,然后再传递到醌池(Q),最后用于NADH的再生或者传递到细胞色素bd-I泛醇氧化酶来还原O2 (图 4)。其中,bc1复合物广泛存在于Zetaproteobacteria铁氧化菌的各个类群,而ACIII仅存在于部分类群,二者功能相同,起到醌还原酶的作用[57]。

|
| 图 4 Zetaproteobacteria铁氧化菌介导Fe2+氧化的电子传递途径模型[57] Figure 4 Electron transport pathway model of Zetaproteobacteria mediated Fe2+ oxidation[57]. Components that are conserved in all Zetaproteobacteria are outlined with a thick line, and components of the pathway from the same module have the same color. |
3.2 铁氧化菌的碳氮硫耦合代谢潜能 3.2.1 固碳途径
Zetaproteobacteria铁氧化菌主要依赖于Fe2+氧化来获取维系自身代谢所需的能量,目前该类铁氧化菌已知的纯培养菌株均为化能自养型微生物。它们的基因组中均具有核酮糖-1, 5-二磷酸羧化酶加氧酶(RuBisCO)基因,表明Calvin-Benson-Bassham (CBB)循环是Zetaproteobacteria铁氧化菌通用的固碳途径(图 5A)[13–14, 64, 70]。RuBisCO基因可以分为Ⅰ和Ⅱ两种类型,其中Ⅰ型RuBisCO对O2不敏感,可以适应高O2浓度的环境;相比之下,Ⅱ型RuBisCO更适应于高CO2和低O2浓度的环境。已有的研究表明,大多数已发现的Zetaproteobacteria铁氧化菌具有Ⅱ型RuBisCO基因,而拥有Ⅰ型RuBisCO基因的Zetaproteobacteria铁氧化菌一般同时也拥有Ⅱ型RuBisCO基因,这意味着Ⅱ型RuBisCO在Zetaproteobacteria铁氧化菌中占主导地位,反映了多数铁氧化菌对低O2浓度环境的偏好与适应(图 5A)。部分Zetaproteobacteria铁氧化菌所同时具有的Ⅰ型RuBisCO基因可以确保它们在O2浓度发生较大波动的情况下仍然能够存活,并保持化学自养的生活方式[13, 29, 43, 64, 66]。

|
| 图 5 Zetaproteobacteria铁氧化菌固碳与氮代谢途径示意图 Figure 5 Schematic diagram of carbon fixation and nitrogen metabolism pathways of Zetaproteobacteria. A: modified after reference [72], most Zetaproteobacteria only encode Form Ⅱ RuBisCO, while Zetaproteobacteria encoding Form Ⅰ RuBisCO often encode both Form Ⅱ. B: all the genes were only found in part of Zetaproteobacteria genomes; green: denitrification; blue: assimilation; purple: nitrogen fixation. |
一个十分有意思的现象是,在部分Zetaproteobacteria铁氧化菌的基因组中还发现了编码丙酮酸甲酸裂解酶(Pfl)、丙酮酸铁氧还蛋白氧化还原酶(Pfor)、磷酸转乙酰酶(Pta)和乙酸激酶(AckA)等与有机质厌氧发酵密切相关的功能基因[71]。但目前为止所有分离得到的Zetaproteobacteria纯培养菌株均被证明严格需氧,所以科学家们推测上述潜在的有机质厌氧发酵代谢途径可能属于“辅助厌氧代谢”的范畴,其产生的能量用于极端条件下短暂维持FeOB细胞的生命而不是用于细胞的生长、发育,从而使得Zetaproteobacteria铁氧化菌在O2波动较大的环境中具备生存优势[71]。
3.2.2 参与氮循环的代谢潜能Zetaproteobacteria铁氧化菌的基因组中还发现了多个与氮循环密切相关的基因。编码同化硝酸盐还原酶(nasAB)、同化亚硝酸盐还原酶(nirBD)、异化硝酸盐还原酶(napAB)、亚硝酸盐还原酶(nirK/nirS)、一氧化氮还原酶(eNOR)以及一氧化二氮还原酶(nosZ)等基因的发现表明,Zetaproteobacteria具有同化氮以及反硝化作用的潜能(图 5B)[36, 52, 64]。理论上,依赖于硝酸盐还原的Fe2+氧化是可行的,并已在可培养的Ferroglobus placidus菌株中得到了验证[52, 73]。Zetaproteobacteria铁氧化菌在进行Fe2+氧化时消耗氧气能够形成厌/极低氧微环境,从而为硝酸盐还原创造有利条件。然而,目前为止,所有已获取的Zetaproteobacteria铁氧化菌的基因组都不具备完整的反硝化途径所需的功能基因,意味着它们不能够独立完成反硝化作用,而是需要与其他微生物共生/共营养的方式来完成这一功能[52]。近期的一项研究证实了上述猜测/假设,在对Lō’ihi海山热液喷口微生物菌席的调查中发现,NO3–通过反硝化途径被还原为N2这一反硝化过程的所有基因在该菌席样品中均被表达。然而,进一步的分析却发现不同反应步骤所对应的不同功能基因却由不同的微生物类群所表达,如在有的样品中Zetaproteobacteria铁氧化菌仅表达了nirK和nosZ两个基因,有的样品中Zetaproteobacteria仅表达eNOR[52]。基于转录组的分析揭示了Zetaproteobacteria铁氧化菌可能是与一种尚未获得分离培养的厌氧异养铁氧化菌Candidatus Ferristratum sp.协同完成了反硝化过程[52]。C. Ferristratum sp.可以同时进行硝酸盐还原与Fe2+氧化,该反应所产生的大量亚硝酸盐被Zetaproteobacteria铁氧化菌所还原。这是一种全新的氮/铁循环耦合模式,厌氧异养铁氧化细菌与微需氧自养铁氧化菌共存并相互依赖,合作完成反硝化反应[52]。
编码铵转运蛋白的amt基因也普遍存在于Zetaproteobacteria铁氧化菌的基因组中,它主要负责介导铁氧化菌对铵的同化过程,表明这些铁氧化微生物更倾向于从周围环境中获取生物可利用的含氮化合物。这也与海底热液喷口及其环境中富含NH4+/NO3–等含氮离子相一致[52, 64, 74–75]。此外,在M. ferrooxydans M34、Mariprofundus sp. EKF-M39和M. ferrooxydans O-1等纯培养铁氧化菌的基因组中还发现了生物固氮所需的nif基因[34, 36, 41],然而其固氮的生物学功能暂未得到证实。
3.2.3 参与硫循环与砷循环的潜能Zetaproteobacteria铁氧化菌的基因组中还发现了编码硫氧化蛋白的基因,如编码硫醌氧化还原酶的sqr基因[27, 32, 40, 75]。硫醌氧化还原酶属于外周膜蛋白,能够催化硫化氢的氧化和泛醌的还原,在嗜酸性氧化亚铁硫杆菌中普遍存在[76]。编码硫双加氧酶的sdo基因在Zetaproteobacteria铁氧化菌的基因组中也有发现。硫双加氧酶是执行单质硫氧化的关键酶之一,属于细胞周质酶,能够在还原型谷胱甘肽存在的前提下将单质硫氧化为亚硫酸盐[27, 77]。
另外,最近的一项研究还从Zetaproteobacteria铁氧化菌的基因组中发现了与元素砷循环密切相关的砷脱毒基因arsC。该基因编码砷酸还原酶,能够将砷酸盐还原成亚砷酸盐,然后通过特异转运蛋白从细胞中排出,从而避免细胞砷中毒[64]。不过,该基因目前仅在一个富砷的热液喷口中被发现,可能反映了Zetaproteobacteria铁氧化菌对特异环境的适应性,并不一定具有普遍性。
需要指出的是,上述与氮、硫和砷相关的代谢潜能证据很多来自于铁氧化菌基因组的功能推测,还需要结合铁氧化菌的分离、培养以及进一步的地球化学培养实验进行验证[52]。
4 结论与展望嗜中性微需氧铁氧化菌是海底热液喷口及其周围环境生态系统的初级生产者之一,其所具有的Fe2+氧化能力对海底岩石的蚀变、热液硫化物的氧化性风化以及低温热液富铁氧化物的沉淀等地质和地球化学过程具有重要影响。随着研究的深入,特别是分离培养技术的突破,新的铁氧化菌不断地被分离培养,高通量测序技术的发展也使得人们获得了大量铁氧化菌的SAGs (single-amplified genomes)与MAGs (metagenome-assembled genomes),结合基因鉴定与功能预测,从而使人们对嗜中性微需氧铁氧化菌的种群多样性、生理特性与代谢潜能以及它们在生态系统中的生态学功能等方面的认识有了巨大的提高。尽管如此,针对海底热液喷口铁氧化菌的研究仍然存在以下几个方面的不足:
(1) 嗜中性微需氧铁氧化菌的生物地理分区:目前,对海底热液系统FeOB的研究主要集中于太平洋海域,特别是夏威夷Lō’ihi海山、马里亚纳弧后热液系统、劳盆地和Juan de Fuca洋脊等。相比之下,聚焦于大西洋和印度洋的研究很少,这些海域栖息的Zetaproteobacteria铁氧化菌的多样性、生理特性,甚至其矿化形成的富铁显微结构的形态等,是否存在地理区域范围内的独特性?目前还知之甚少,需要有针对性地开展对比研究。
(2) 铁氧化菌生命活动形成的显微结构与铁氧化种群之间的对应关系:尽管一些具有独特形貌特征的显微结构可以作为FeOB存在的判别标志,然而并没有建立显微结构与特定FeOB类群之间的一一对应关系。而且,越来越多的研究发现了大量与前文所述FeOB所产生的典型形貌所不同的显微结构,可能意味着新的铁氧化种群的存在。
(3) Fe2+氧化过程与其他元素地球化学循环过程的耦合研究:前文提到了Zetaproteobacteria铁氧化菌还具备与氮、硫、氢以及砷元素循环相关的功能基因,然而上述基因的生物学功能还没有得到证实。需要我们进一步开展铁氧化菌的分离和培养以获得新的Zetaproteobacteria纯菌,进而开展地球化学培养实验,同时结合宏转录组和蛋白质组学的研究,从而为铁氧化菌的上述代谢功能提供直接证据。
(4) 铁氧化菌与条带状含铁建造:前寒武纪(主要为太古宙到古元古代)条带状含铁建造(BIF)的成因一直有争议,微需氧铁氧化菌介导的铁氧化是人们提出的成因之一。基于可培养嗜中性微需氧FeOB纯培养的模拟实验可以帮助我们回答地球早期BIF形成的一系列问题。
完善上述研究对于进一步认识和拓展海底热液系统关键成矿元素的迁移与富集规律、喷口生态系统以及生物与极端环境的相互作用机制、微生物生物成矿理论等方面具有重要的理论价值,同时对寻求地球早期生命活动的显微地质记录与化石标志、地外生命的探索与发现、丰富地球生命形式与物种多样性、揭示生命能量代谢新途径等方面具有重要的参考价值。
References
| [1] | Frey PA, Reed GH. The ubiquity of iron. ACS Chemical Biology, 2012, 7(9): 1477-1481. DOI:10.1021/cb300323q |
| [2] | Hedrich S, Schlömann M, Johnson DB. The iron-oxidizing proteobacteria. Microbiology: Reading, England, 2011, 157(Pt 6): 1551-1564. |
| [3] | Melton ED, Swanner ED, Behrens S, Schmidt C, Kappler A. The interplay of microbially mediated and abiotic reactions in the biogeochemical Fe cycle. Nature Reviews Microbiology, 2014, 12(12): 797-808. DOI:10.1038/nrmicro3347 |
| [4] | Druschel GK, Emerson D, Sutka R, Suchecki P, Luther GW Ⅲ. Low-oxygen and chemical kinetic constraints on the geochemical niche of neutrophilic iron(Ⅱ) oxidizing microorganisms. Geochimica et Cosmochimica Acta, 2008, 72(14): 3358-3370. DOI:10.1016/j.gca.2008.04.035 |
| [5] | Emerson D, Fleming EJ, McBeth JM. Iron-oxidizing bacteria: an environmental and genomic perspective. Annual Review of Microbiology, 2010, 64: 561-583. DOI:10.1146/annurev.micro.112408.134208 |
| [6] | Lueder U, Druschel G, Emerson D, Kappler A, Schmidt C. Quantitative analysis of O2 and Fe2+ profiles in gradient tubes for cultivation of microaerophilic Iron(Ⅱ)-oxidizing bacteria. FEMS Microbiology Ecology, 2018, 94(2). DOI:10.1093/femsec/fix177 |
| [7] | Maisch M, Lueder U, Laufer K, Scholze C, Kappler A, Schmidt C. Contribution of microaerophilic iron(Ⅱ)-oxidizers to iron(Ⅲ) mineral formation. Environmental Science & Technology, 2019, 53(14): 8197-8204. |
| [8] | Ehrenberg. Vorläufige mittheilungen über das wirkliche vorkommen fossiler infusorien und ihre grosse verbreitung. Annalen Der Physik Und Chemie, 1836, 114(5): 213-227. DOI:10.1002/andp.18361140520 |
| [9] | Laufer K, Nordhoff M, Halama M, Martinez RE, Obst M, Nowak M, Stryhanyuk H, Richnow HH, Kappler A. Microaerophilic Fe(Ⅱ)-oxidizing Zetaproteobacteria isolated from low-Fe marine coastal sediments: physiology and composition of their twisted stalks. Applied and Environmental Microbiology, 2017, 83(8): e03118-16. |
| [10] | Chan CS, McAllister SM, Leavitt AH, Glazer BT, Krepski ST, Emerson D. The architecture of iron microbial mats reflects the adaptation of chemolithotrophic iron oxidation in freshwater and marine environments. Frontiers in Microbiology, 2016, 7: 796. |
| [11] | Chan CS, Fakra SC, Emerson D, Fleming EJ, Edwards KJ. Lithotrophic iron-oxidizing bacteria produce organic stalks to control mineral growth: implications for biosignature formation. The ISME Journal, 2011, 5(4): 717-727. |
| [12] | Kappler A, Bryce C, Mansor M, Lueder U, Byrne JM, Swanner ED. An evolving view on biogeochemical cycling of iron. Nature Reviews Microbiology, 2021, 19(6): 360-374. DOI:10.1038/s41579-020-00502-7 |
| [13] | McAllister SM, Moore RM, Gartman A, Luther GW, Emerson D, Chan CS. The Fe(Ⅱ)-oxidizing Zetaproteobacteria: historical, ecological and genomic perspectives. FEMS Microbiology Ecology, 2019, 95(4): fiz015. |
| [14] | Mori JF, Scott JJ, Hager KW, Moyer CL, Küsel K, Emerson D. Physiological and ecological implications of an iron- or hydrogen-oxidizing member of the Zetaproteobacteria, Ghiorsea bivora, gen. nov., sp. nov.. The ISME Journal, 2017, 11(11): 2624-2636. DOI:10.1038/ismej.2017.132 |
| [15] | Emerson D, Rentz JA, Lilburn TG, Davis RE, Aldrich H, Chan C, Moyer CL. A novel lineage of proteobacteria involved in formation of marine Fe-oxidizing microbial mat communities. PLoS One, 2007, 2(7): e667. |
| [16] | Emerson D, Scott JJ, Leavitt A, Fleming E, Moyer C. In situ estimates of iron-oxidation and accretion rates for iron-oxidizing bacterial mats at Lō'ihi Seamount. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 2017, 126: 31-39. DOI:10.1016/j.dsr.2017.05.011 |
| [17] | Juniper SK, Fouquet Y. Filamentous iron-silica deposits from modern and ancient hydrothermal sites. Canadian Mineralogist, 1988, 26 pt 3: 859-869. |
| [18] | Ehrenberg CG. Die infusionsthierchen als vollkommene organismen. Ein blick in das tiefere organische leben der natur. Leipzig, : L. Voss, 1838. |
| [19] | Kützing FT. Phycologia generalis: oder, Anatomie, physiologie und systemkunde der tange/Bearb. Von Friedrich Traugott Kützing. Mit 80 farbig gedruckten tafeln, gezeichnet und gravirt vom verfasser. Leipzig: F. A. Brockhaus, 1843. |
| [20] | Emerson D, Moyer CL. Neutrophilic Fe-oxidizing bacteria are abundant at the Loihi Seamount hydrothermal vents and play a major role in Fe oxide deposition. Applied and Environmental Microbiology, 2002, 68(6): 3085-3093. DOI:10.1128/AEM.68.6.3085-3093.2002 |
| [21] | Cambon-Bonavita MA, Aubé J, Cueff-Gauchard V, Reveillaud J. Niche partitioning in the Rimicaris exoculata holobiont: the case of the first symbiotic Zetaproteobacteria. Microbiome, 2021, 9(1): 87. DOI:10.1186/s40168-021-01045-6 |
| [22] | Forget NL, Murdock SA, Juniper SK. Bacterial diversity in Fe-rich hydrothermal sediments at two South Tonga Arc submarine volcanoes. Geobiology, 2010, 8(5): 417-432. DOI:10.1111/j.1472-4669.2010.00247.x |
| [23] | Henri PA, Rommevaux-Jestin C, Lesongeur F, Mumford A, Emerson D, Godfroy A, Ménez B. Structural iron (Ⅱ) of basaltic glass as an energy source for Zetaproteobacteria in an abyssal plain environment, off the mid Atlantic ridge. Frontiers in Microbiology, 2016, 6: 1518. |
| [24] | Jan C, Petersen JM, Werner J, Teeling H, Huang SX, Glöckner FO, Golyshina OV, Dubilier N, Golyshin PN, Jebbar M, Cambon-Bonavita MA. The gill chamber epibiosis of deep-sea shrimp Rimicaris exoculata: an in-depth metagenomic investigation and discovery of Zetaproteobacteria. Environmental Microbiology, 2014, 16(9): 2723-2738. DOI:10.1111/1462-2920.12406 |
| [25] | Kato S, Yanagawa K, Sunamura M, Takano Y, Ishibashi JI, Kakegawa T, Utsumi M, Yamanaka T, Toki T, Noguchi T, Kobayashi K, Moroi A, Kimura H, Kawarabayasi Y, Marumo K, Urabe T, Yamagishi A. Abundance of Zetaproteobacteria within crustal fluids in back-arc hydrothermal fields of the Southern Mariana Trough. Environmental Microbiology, 2009, 11(12): 3210-3222. DOI:10.1111/j.1462-2920.2009.02031.x |
| [26] | Li JT, Zhou HY, Peng XT, Wu ZJ, Chen S, Fang JS. Microbial diversity and biomineralization in low-temperature hydrothermal iron-silica-rich precipitates of the Lau Basin hydrothermal field. FEMS Microbiology Ecology, 2012, 81(1): 205-216. DOI:10.1111/j.1574-6941.2012.01367.x |
| [27] | Tully BJ, Wheat CG, Glazer BT, Huber JA. A dynamic microbial community with high functional redundancy inhabits the cold, oxic subseafloor aquifer. The ISME Journal, 2018, 12(1): 1-16. DOI:10.1038/ismej.2017.187 |
| [28] | Beam JP, Scott JJ, McAllister SM, Chan CS, McManus J, Meysman FJR, Emerson D. Biological rejuvenation of iron oxides in bioturbated marine sediments. The ISME Journal, 2018, 12(5): 1389-1394. DOI:10.1038/s41396-017-0032-6 |
| [29] | Chiu BK, Kato S, McAllister SM, Field EK, Chan CS. Novel pelagic iron-oxidizing Zetaproteobacteria from the Chesapeake Bay oxic–anoxic transition zone. Frontiers in Microbiology, 2017, 8: 1280. DOI:10.3389/fmicb.2017.01280 |
| [30] | Emerson JB, Thomas BC, Alvarez W, Banfield JF. Metagenomic analysis of a high carbon dioxide subsurface microbial community populated by chemolithoautotrophs and bacteria and archaea from candidate phyla. Environmental Microbiology, 2016, 18(6): 1686-1703. DOI:10.1111/1462-2920.12817 |
| [31] | Dalcin Martins P, Danczak RE, Roux S, Frank J, Borton MA, Wolfe RA, Burris MN, Wilkins MJ. Viral and metabolic controls on high rates of microbial sulfur and carbon cycling in wetland ecosystems. Microbiome, 2018, 6(1): 138. DOI:10.1186/s40168-018-0522-4 |
| [32] | Probst AJ, Castelle CJ, Singh A, Brown CT, Anantharaman K, Sharon I, Hug LA, Burstein D, Emerson JB, Thomas BC, Banfield JF. Genomic resolution of a cold subsurface aquifer community provides metabolic insights for novel microbes adapted to high CO2 concentrations. Environmental Microbiology, 2017, 19(2): 459-474. DOI:10.1111/1462-2920.13362 |
| [33] | Probst AJ, Ladd B, Jarett JK, Geller-McGrath DE, Sieber CMK, Emerson JB, Anantharaman K, Thomas BC, Malmstrom RR, Stieglmeier M, Klingl A, Woyke T, Ryan MC, Banfield JF. Differential depth distribution of microbial function and putative symbionts through sediment-hosted aquifers in the deep terrestrial subsurface. Nature Microbiology, 2018, 3(3): 328-336. DOI:10.1038/s41564-017-0098-y |
| [34] | McAllister SM, Davis RE, McBeth JM, Tebo BM, Emerson D, Moyer CL. Biodiversity and emerging biogeography of the neutrophilic iron-oxidizing Zetaproteobacteria. Applied and Environmental Microbiology, 2011, 77(15): 5445-5457. DOI:10.1128/AEM.00533-11 |
| [35] | Emerson D, Merrill Floyd M. Enrichment and isolation of iron-oxidizing bacteria at neutral pH. Methods in Enzymology, 2005, 397: 112-123. |
| [36] | Field EK, Sczyrba A, Lyman AE, Harris CC, Woyke T, Stepanauskas R, Emerson D. Genomic insights into the uncultivated marine Zetaproteobacteria at Lo'ihi Seamount. The ISME Journal, 2015, 9(4): 857-870. DOI:10.1038/ismej.2014.183 |
| [37] | Garrison CE, Price KA, Field EK. Environmental evidence for and genomic insight into the preference of iron-oxidizing bacteria for more-corrosion-resistant stainless steel at higher salinities. Applied and Environmental Microbiology, 2019, 85(14): e00483-e00419. |
| [38] | Koeksoy E, Bezuidt OM, Bayer T, Chan CS, Emerson D. Zetaproteobacteria pan-genome reveals candidate gene cluster for twisted stalk biosynthesis and export. Frontiers in Microbiology, 2021, 12: 679409. DOI:10.3389/fmicb.2021.679409 |
| [39] | Makita H, Tanaka E, Mitsunobu S, Miyazaki M, Nunoura T, Uematsu K, Takaki Y, Nishi S, Shimamura S, Ken TK. Mariprofundus micogutta sp. nov., a novel iron-oxidizing Zetaproteobacterium isolated from a deep-sea hydrothermal field at the Bayonnaise knoll of the Izu-Ogasawara arc, and a description of Mariprofundales ord. nov. and Zetaproteobacteria classis nov. Archives of Microbiology, 2017, 199(2): 335-346. DOI:10.1007/s00203-016-1307-4 |
| [40] | Lopez A, Albino D, Beraki S, Broomell S, Canela R, Dingmon T, Estrada S, Fernandez M, Savalia P, Nealson K, Emerson D, Barco R, Tully BJ, Amend JP. Genome sequence of Mariprofundus sp. strain EBB-1, a novel marine autotroph isolated from an iron-sulfur mineral. Microbiology Resource Announcements, 2019, 8(39): e00995-19. |
| [41] | Price KA, Garrison CE, Richards N, Field EK. A shallow water ferrous-hulled shipwreck reveals a distinct microbial community. Frontiers in Microbiology, 2020, 11: 1897. DOI:10.3389/fmicb.2020.01897 |
| [42] | Moreira APB, Meirelles PM, Thompson F. The family Mariprofundaceae//Rosenberg E, DeLong EF, Lory S, Stackebrandt E, Thompson F (eds). The Prokaryotes. Berlin: Springer, 2014. |
| [43] | Blackwell N, Bryce C, Straub D, Kappler A, Kleindienst S. Genomic insights into two novel Fe(Ⅱ)-oxidizing Zetaproteobacteria isolates reveal lifestyle adaption to coastal marine sediments. Applied and Environmental Microbiology, 2020, 86(17): e01660-20. |
| [44] | Duchinski K, Moyer CL, Hager K, Fullerton H. Fine-scale biogeography and the inference of ecological interactions among neutrophilic iron-oxidizing Zetaproteobacteria as determined by a rule-based microbial network. Frontiers in Microbiology, 2019, 10: 2389. DOI:10.3389/fmicb.2019.02389 |
| [45] | Edwards KJ, Rogers DR, Wirsen CO, McCollom TM. Isolation and characterization of novel psychrophilic, neutrophilic, Fe-oxidizing, chemolithoautotrophic Alpha- and Gamma-proteobacteria from the deep sea. Applied and Environmental Microbiology, 2003, 69(5): 2906-2913. DOI:10.1128/AEM.69.5.2906-2913.2003 |
| [46] | Sudek LA, Templeton AS, Tebo BM, Staudigel H. Microbial ecology of Fe (hydr)oxide mats and basaltic rock from vailulu'u seamount, American Samoa. Geomicrobiology Journal, 2009, 26(8): 581-596. DOI:10.1080/01490450903263400 |
| [47] | Li JT, Cui JM, Yang QH, Cui GJ, Wei BB, Wu ZJ, Wang Y, Zhou HY. Oxidative weathering and microbial diversity of an inactive seafloor hydrothermal sulfide chimney. Frontiers in Microbiology, 2017, 8: 1378. DOI:10.3389/fmicb.2017.01378 |
| [48] | Meier DV, Pjevac P, Bach W, Markert S, Schweder T, Jamieson J, Petersen S, Amann R, Meyerdierks A. Microbial metal-sulfide oxidation in inactive hydrothermal vent chimneys suggested by metagenomic and metaproteomic analyses. Environmental Microbiology, 2019, 21(2): 682-701. DOI:10.1111/1462-2920.14514 |
| [49] | Meyer JL, Akerman NH, Proskurowski G, Huber JA. Microbiological characterization of post-eruption "snowblower" vents at axial seamount, Juan de Fuca ridge. Frontiers in Microbiology, 2013, 4: 153. |
| [50] | Sylvan JB, Pyenson BC, Rouxel O, German CR, Edwards KJ. Time-series analysis of two hydrothermal plumes at 9°50'N East Pacific Rise reveals distinct, heterogeneous bacterial populations. Geobiology, 2012, 10(2): 178-192. DOI:10.1111/j.1472-4669.2011.00315.x |
| [51] | Vander Roost J, Daae FL, Steen IH, Thorseth IH, Dahle H. Distribution patterns of iron-oxidizing Zeta- and Beta- proteobacteria from different environmental settings at the jan Mayen vent fields. Frontiers in Microbiology, 2018, 9: 3008. DOI:10.3389/fmicb.2018.03008 |
| [52] | McAllister SM, Vandzura R, Keffer JL, Polson SW, Chan CS. Aerobic and anaerobic iron oxidizers together drive denitrification and carbon cycling at marine iron-rich hydrothermal vents. The ISME Journal, 2021, 15(5): 1271-1286. DOI:10.1038/s41396-020-00849-y |
| [53] | Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution, 2018, 35(6): 1547-1549. DOI:10.1093/molbev/msy096 |
| [54] | McBeth JM, Little BJ, Ray RI, Farrar KM, Emerson D. Neutrophilic iron-oxidizing "Zetaproteobacteria" and mild steel corrosion in nearshore marine environments. Applied and Environmental Microbiology, 2011, 77(4): 1405-1412. DOI:10.1128/AEM.02095-10 |
| [55] | Barco RA, Hoffman CL, Ramírez GA, Toner BM, Edwards KJ, Sylvan JB. In-situ incubation of iron-sulfur mineral reveals a diverse chemolithoautotrophic community and a new biogeochemical role for Thiomicrospira. Environmental Microbiology, 2017, 19(3): 1322-1337. DOI:10.1111/1462-2920.13666 |
| [56] | Mumford AC, Adaktylou IJ, Emerson D. Peeking under the iron curtain: development of a microcosm for imaging the colonization of steel surfaces by Mariprofundus sp. strain DIS-1, an oxygen-tolerant Fe-oxidizing bacterium. Applied and Environmental Microbiology, 2016, 82(22): 6799-6807. |
| [57] | McAllister SM, Polson SW, Butterfield DA, Glazer BT, Sylvan JB, Chan CS. Validating the Cyc2 neutrophilic iron oxidation pathway using meta-omics of Zetaproteobacteria iron mats at marine hydrothermal vents. mSystems, 2020, 5(1): e00553-e00519. |
| [58] | Byrne JM, Schmidt M, Gauger T, Bryce C, Kappler A. Imaging organic–mineral aggregates formed by Fe(Ⅱ)-oxidizing bacteria using helium ion microscopy. Environmental Science & Technology Letters, 2018, 5(4): 209-213. |
| [59] | Li JT, Su L, Wang F, Yang JY, Gu LY, Sun MX, Li QR, Zhou HY, Fang JS. Elucidating the biomineralization of low-temperature hydrothermal precipitates with varying Fe, Si contents: indication from ultrastructure and microbiological analyses. Deep Sea Research Part I: Oceanographic Research Papers, 2020, 157: 103208. DOI:10.1016/j.dsr.2019.103208 |
| [60] | Edwards KJ, Wheat CG, Sylvan JB. Under the sea: microbial life in volcanic oceanic crust. Nature Reviews Microbiology, 2011, 9(10): 703-712. DOI:10.1038/nrmicro2647 |
| [61] | Johannessen KC, Vander Roost J, Dahle H, Dundas SH, Pedersen RB, Thorseth IH. Environmental controls on biomineralization and Fe-mound formation in a low-temperature hydrothermal system at the Jan Mayen Vent Fields. Geochimica et Cosmochimica Acta, 2017, 202: 101-123. DOI:10.1016/j.gca.2016.12.016 |
| [62] | Krepski ST, Emerson D, Hredzak-Showalter PL, Luther GWI, Chan CS. Morphology of biogenic iron oxides records microbial physiology and environmental conditions: toward interpreting iron microfossils. Geobiology, 2013, 11(5): 457-471. DOI:10.1111/gbi.12043 |
| [63] | Fleming EJ, Davis RE, McAllister SM, Chan CS, Moyer CL, Tebo BM, Emerson D. Hidden in plain sight: discovery of sheath-forming, iron-oxidizing Zetaproteobacteria at Loihi Seamount, Hawaii, USA. FEMS Microbiology Ecology, 2013, 85(1): 116-127. DOI:10.1111/1574-6941.12104 |
| [64] | Fullerton H, Hager KW, McAllister SM, Moyer CL. Hidden diversity revealed by genome-resolved metagenomics of iron-oxidizing microbial mats from Lō'ihi Seamount, Hawai'i. The ISME Journal, 2017, 11(8): 1900-1914. DOI:10.1038/ismej.2017.40 |
| [65] | Makita H. Iron-oxidizing bacteria in marine environments: recent progresses and future directions. World Journal of Microbiology & Biotechnology, 2018, 34(8): 110. |
| [66] | Barco RA, Emerson D, Sylvan JB, Orcutt BN, Jacobson Meyers ME, Ramírez GA, Zhong JD, Edwards KJ. New insight into microbial iron oxidation as revealed by the proteomic profile of an obligate iron-oxidizing chemolithoautotroph. Applied and Environmental Microbiology, 2015, 81(17): 5927-5937. DOI:10.1128/AEM.01374-15 |
| [67] | Keffer JL, McAllister SM, Garber A, Hallahan BJ, Sutherland MC, Rozovsky S, Chan CS. Iron oxidation by a fused cytochrome-porin common to diverse iron-oxidizing bacteria. bioRxiv, 2021. DOI:10.1101/228056 |
| [68] | Castelle C, Guiral M, Malarte G, Ledgham F, Leroy G, Brugna M, Giudici-Orticoni MT. A new iron-oxidizing/O2– reducing super complex spanning both inner and outer membranes, isolated from the extreme acidophile Acidithiobacillus ferrooxidans. Journal of Biological Chemistry, 2008, 283(38): 25803-25811. DOI:10.1074/jbc.M802496200 |
| [69] | Emerson D, Field EK, Chertkov O, Davenport KW, Goodwin L, Munk C, Nolan M, Woyke T. Comparative genomics of freshwater Fe-oxidizing bacteria: implications for physiology, ecology, and systematics. Frontiers in Microbiology, 2013, 4: 254. |
| [70] | Singer E, Emerson D, Webb EA, Barco RA, Kuenen JG, Nelson WC, Chan CS, Comolli LR, Ferriera S, Johnson J, Heidelberg JF, Edwards KJ. Mariprofundus ferrooxydans PV-1 the first genome of a marine Fe(Ⅱ) oxidizing Zetaproteobacterium. PLoS One, 2011, 6(9): e25386. DOI:10.1371/journal.pone.0025386 |
| [71] | Jain A, Gralnick JA. Evidence for auxiliary anaerobic metabolism in obligately aerobic Zetaproteobacteria. The ISME Journal, 2020, 14(4): 1057-1062. DOI:10.1038/s41396-020-0586-6 |
| [72] |
Zhu SG, Xu CF. Biochemistry. 4th Edition. Beijing: Higher Education Press, 2016. (in Chinese) 朱圣庚, 徐长法. 生物化学. 第四版. 北京: 高等教育出版社, 2016. |
| [73] | Hafenbradl D, Keller M, Dirmeier R, Rachel R, Roßnagel P, Burggraf S, Huber H, Stetter KO. Ferroglobus placidus gen. nov., sp. nov., a novel hyperthermophilic archaeum that oxidizes Fe2+ at neutral pH under anoxic conditions. Archives of Microbiology, 1996, 166(5): 308-314. |
| [74] | Jesser KJ, Fullerton H, Hager KW, Moyer CL. Quantitative PCR analysis of functional genes in iron-rich microbial mats at an active hydrothermal vent system (Lō'ihi Seamount, Hawai'i). Applied and Environmental Microbiology, 2015, 81(9): 2976-2984. DOI:10.1128/AEM.03608-14 |
| [75] | Singer E, Heidelberg JF, Dhillon A, Edwards KJ. Metagenomic insights into the dominant Fe(Ⅱ) oxidizing Zetaproteobacteria from an iron mat at Lō'ihi, Hawai'l. Frontiers in Microbiology, 2013, 4: 52. |
| [76] | Zhang YF, Qadri A, Weiner JH. The quinone-binding site of Acidithiobacillus ferrooxidans sulfide: quinone oxidoreductase controls both sulfide oxidation and quinone reduction. Biochemistry and Cell Biology, 2016, 94(2): 159-166. DOI:10.1139/bcb-2015-0097 |
| [77] | Wang R, Lin JQ, Liu XM, Pang X, Zhang CJ, Yang CL, Gao XY, Lin CM, Li YQ, Li Y, Lin JQ, Chen LX. Sulfur oxidation in the acidophilic autotrophic Acidithiobacillus spp. Frontiers in Microbiology, 2019, 9: 3290. DOI:10.3389/fmicb.2018.03290 |