 2023, Vol. 63
2023, Vol. 63中国科学院微生物研究所,中国微生物学会
文章信息
- 鲁妍璇, 曹毅, 李博雅, 吕朝阳, 谢家贝, 张克诚, 葛蓓孛, 冉隆贤. 2023
- LU Yanxuan, CAO Yi, LI Boya, LÜ Zhaoyang, XIE Jiabei, ZHANG Kecheng, GE Beibei, RAN Longxian.
- 利迪链霉菌K2对灰霉病菌的抑菌效果及抑菌物质鉴定
- Inhibition of Streptomyces lydicus K2 on Botrytis cinerea and identification of its inhibitory substances
- 微生物学报, 63(5): 1991-2006
- Acta Microbiologica Sinica, 63(5): 1991-2006
-
文章历史
- 收稿日期:2023-02-28


引用本文 |

 ,
, 2. 贵州省烟草科学研究院, 贵州 贵阳 550081;
3. 中国农业科学院植物保护研究所 植物病虫害综合治理全国重点实验室, 北京 100193
2. Guizhou Academy of Tobacco Science, Guiyang 550081, Guizhou, China;
3. State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China
灰霉病是由灰葡萄孢(Botrytis cinerea)引起的可危害蔬菜、果树及花卉等多种植物的重要病害,在果蔬生产中的危害尤为严重[1],并在果实贮藏运输中加快消耗贮藏物的营养物质,致使贮藏物的储存期变短、品质下降而引起严重的经济损失[2]。现阶段,对果蔬生产中及采后储藏期灰霉病主要使用化学杀菌剂进行防治,其虽具有高效的特点,但由于灰霉病菌寄主范围广、繁殖快、遗传变异大,易产生抗药性[3],并且化学药剂的残留对环境造成了不可逆的污染,危及人畜健康。
生物防治因其具有持效期长和环境友好等优点而受到广泛的关注,筛选出高效多功能的生防微生物是目前的研究热点之一[4]。在植物病害生防菌中,链霉菌在代谢过程中能产生多种抗生素等次生代谢物,这些代谢物具有高效、低毒的特点。如武夷菌素是由不吸水链霉菌武夷变种CK-15产生的一种广谱、高效、低毒的农用抗生素,生产上对草莓灰霉病、番茄灰霉病具有较好的田间防治效果[5]。研究还发现微生物能利用大分子物质如蛋白质、脂肪和碳水化合物等生成葡萄糖、氨基酸和脂肪酸等小分子代谢产物,再进行初级代谢和次级代谢将这些物质转化能产生大量种类丰富、功能多样的挥发性物质(volatile organic compounds, VOCs)[6]。微生物VOCs成分复杂、功能多样,据报道微生物VOCs具有极强的抑菌效果,能够协同作用,抑制甚至杀死果蔬采后病原菌,有些还能促进植物生长和作物增产,而且它们具有在空气、土壤空隙中高效率的扩散能力,能快速扩大作用范围[7]。在果实病害防治中,挥发性物质可以不与果实直接接触,也不会在果实表面留下残留物[8],从而可以减少化肥和农药的使用,且有利于保障种植和采后贮藏环节的农产品质量安全。因此对链霉菌非挥发和挥发性代谢产物进行开发对植物病害防治以及采后果实保鲜等方面具有广泛的研究价值和应用前景。
为了减轻或避免灰霉病菌在作物生产及产品储藏期间的危害,本研究从土壤中筛选到一株对深红酵母具有强拮抗作用的链霉菌K2。通过16S rRNA基因对该菌株进行初步鉴定,同时测定其培养液及其挥发性物质对灰霉病菌的抑菌效果,并对其培养液中的活性成分及产挥发性成分进行检测及鉴定,为链霉菌K2作为防治灰霉病新型生物农药的开发提供科学依据和理论支持。
1 材料与方法 1.1 供试材料菌株K2从土壤样品中分离、纯化获得;菌株CK-15,用于活性比对;供试病原真菌及细菌:灰霉病菌、苹果腐烂病菌、苹果轮纹病菌、核盘菌、棉花枯萎病菌、烟草赤星病菌、杨树溃疡病菌、小麦纹枯病菌、辣椒疫病病菌、玉米大斑病菌、西瓜果斑病菌、辣椒疮痂病菌、青枯病菌、黄瓜角斑病菌、水稻白叶枯病菌和深红酵母均由中国农业科学院植物保护研究所农用抗生素组保存,用于抑菌活性检测。
PDA培养基:去皮马铃薯200 g,葡萄糖20 g,琼脂粉15 g,蒸馏水1 000 mL,pH 7.2。PDA培养基用于病原真菌的培养;
MS培养基:20 g黄豆饼放入蒸馏水中煮30 min,用4层纱布过滤定容至1 000 mL,每100 mL分装入盛有2 g甘露醇和1.7 g琼脂粉的250 mL三角瓶中。MS培养基用于菌株CK-15的培养;
LB培养基:胰蛋白胨10 g,酵母提取物5 g,氯化钠10 g,琼脂粉15 g,蒸馏水1 000 mL,pH 7.2。LB培养基用于病原细菌的培养;
ISP2培养基:酵母膏10 g,琼脂粉20 g,麦芽浸粉10 g,葡萄糖4 g,琼脂粉20 g,蒸馏水1 000 mL,pH 7.2–7.4。ISP2培养基用于菌株K2的培养;
种子培养基:葡萄糖20 g,蛋白胨6 g,酵母粉6 g,氯化钠10 g,蒸馏水1 000 mL;
液体培养基A:黄豆粉20 g,玉米粉30 g,葡萄糖20 g,硫酸铵4 g,碳酸钙3 g,蒸馏水1 000 mL,pH 7.2–7.4。菌株CK-15的最适发酵培养基;
液体培养基B:黄豆粉15 g,可溶性淀粉20 g,酵母粉5 g,蛋白胨2 g,氯化钠4 g,碳酸钙4 g,蒸馏水1 000 mL,pH 7.2–7.4;
麦粒培养基:用清水浸泡小麦粒24 h,再放到沸水中煮至小麦粒表皮裂开,过滤晾干后按每瓶20 g分装到250 mL三角瓶,加入质量比为1:1的蒸馏水。
上述各种培养基均于121 ℃灭菌20 min后备用。
1.2 灭菌培养液的抗菌活性检测培养液的制备:菌株K2划线接种在ISP2培养基平板上,菌株CK-15划线接种在MS培养基平板上,均放置于28 ℃培养箱中培养7 d,将培养好的菌株切成1.0 cm2大小后接入种子培养基中,置于28 ℃、220 r/min振荡培养24 h,用紫外分光光度测定其OD600,以1%的接种量接种于液体培养基A、液体培养基B中,菌株CK-15重复上述操作只接于液体培养基A中,液体培养基在28 ℃、220 r/min条件下振荡培养72 h,将培养液用滤纸过滤后再在无菌条件下用0.22 μm微孔滤膜过滤除菌,将无菌培养液于4 ℃保存备用。
病原真菌拮抗能力测定:将培养好的病原真菌用打孔器打取直径为5 mm的菌块,培养皿中倒入PDA培养基,在培养皿中心放置打取好的病原真菌菌块,在距离中心30 mm角度120 °设置三点分别放置1个牛津杯,牛津杯中分别接菌株培养液A、培养液B和CK-15培养液A,培养液的量为200 μL,每种培养液重复3次。
病原细菌拮抗能力测定:将病原细菌挑取至LB液体培养基,于28 ℃、180 r/min振荡培养24 h,将摇瓶培养细菌稀释,分光光度计测定OD600为0.1–0.2,取稀释后的细菌菌液200 μL均匀涂布至LA固体培养基上,距离培养皿中心30 mm角度120 °设置3个点,在每点处分别接菌株培养液A、培养液B和CK-15培养液A,培养液液量为10 μL。每种培养液重复3次。
深红酵母拮抗能力测定:深红酵母先用无菌水洗脱,待PDA培养基温度至45 ℃,吸取1 mL红酵母至PDA培养基摇匀。距离培养皿中心30 mm角度120 °设置三点分别放置1个牛津杯,牛津杯中分别接菌株培养液A、培养液B和CK-15培养液A,培养液的量为200 μL,每种培养液重复3次。
抑菌作用测定:上述各种处理后,在28 ℃恒温箱中黑暗培养5 d,取出测量抑菌带的大小,由此确定3类菌对灰霉菌的拮抗效果:抑菌带宽 < 13.8 mm时,抑菌作用弱;抑菌带宽≥13.8 mm时,抑菌作用强。
1.3 K2产挥发性物质对灰霉病菌的抑菌活性测定链霉菌麦粒培养基的制备:菌株K2划线接种在ISP2培养基平板上,菌株CK-15划线接种在MS培养基平板上,均放置于28 ℃培养箱中培养7 d,将培养好的菌株切成1.0 cm2大小后接入种子培养基中,置于28 ℃、220 r/min振荡培养24 h,用紫外分光光度测定其OD600,以20%的接种量接于麦粒培养基上,混匀后放到温度为28 ℃的培养箱中静置培养,每天摇瓶2次以使链霉菌在麦粒培养基上均匀生长,培养10 d至麦粒表面密布灰白色菌落即得到链霉菌麦粒培养基备用。
选用2种测定方法进行K2菌株产挥发性物质对灰霉病菌的抑制效果测定。采用双皿对峙法[9],将培养好的病原真菌用打孔器打取直径为5 mm的菌块,取2个大小为90 mm培养皿,一个倒入PDA培养基,冷却后将直径为5 mm灰霉病菌菌块放置于培养基中央。再取已制备好的链霉菌麦粒培养物,放入另一个皿底中,将有灰霉病菌菌块接种含麦粒培养基上,以添加等量未接种链霉菌种子液的麦粒培养基为对照,用封口膜封口。
参照李其利[10]的方法,将培养好的病原真菌用打孔器打取直径为5 mm的菌块,在直径180 mm培养皿中放置2个90 mm培养皿,在培养基中央接灰霉病菌菌块,另一个70 mm培养皿中放置已制备好的链霉菌麦粒培养基,以添加等量未接种链霉菌种子液的麦粒培养基为对照,然后将大培养皿密封。
将以上处理后封口的培养皿置于25 ℃恒温培养箱中培养,5 d后采用十字交叉法测量处理组和对照组菌落直径。每个处理重复3次。测量得到菌落直径,根据公式(1)计算挥发性物质的抑菌率。
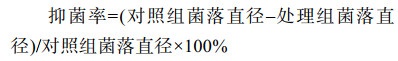
|
公式(1) |
采用16S rRNA基因技术对菌株进行种属鉴定。菌株DNA的提取使用链霉菌全基因组DNA提取试剂盒。16S rRNA基因通用引物27F/1492R (AGAGTTTGATCMTGGCTCAG/GGTTACCTTGTTACGACTT)由华大基因合成。PCR扩增的反应体系(50 μL)含2×Taq PCR Master Mix预混合液25 μL,27F 2 μL,1492R 2 μL,DNA模板1 μL及ddH2O 20 μL;PCR反应条件设置为:95 ℃预变性5 min;95 ℃变性30 s,45 ℃退火15 s,72 ℃延伸30 s,循环35次;最后72 ℃延伸10 min。PCR扩增结束后,将PCR产物用1%琼脂糖凝胶进行电泳;将PCR扩增产物送北京擎科生物技术有限公司测序,测序结果提交至NCBI中进行序列比对,选择与K2同源性较高的菌株,应用MEGA 7.0软件进行多序列比对并建立系统发育树。
1.5 K2培养液中活性成分定性分析标准品溶液配制:准确称取各标准品2 mg溶于2 mL蒸馏水中,充分混匀后即得到1 000 μg/mL标准溶液,液相色谱-质谱检测中标准品需稀释至1 μg/mL。取1.5 mL标准品溶液及无菌培养液分别经0.22 μm的无菌滤膜过滤后置于液相进样瓶中待检测;流动相均需在超声波清洗机中超声波处理30 min。将标准品溶液和样品溶液在同一条件下进行检测,通过分析比较标准品与样品色谱图出峰保留时间进行定性分析。
高效液相色谱(high performance liquid chromatography, HPLC)检测条件(表 1);液质色谱-质谱检测中液相色谱条件:柱温35 ℃,进样量2 μL,流速0.3 mL/min,流动相A为甲醇,B为0.1%甲酸水溶液,梯度洗脱程序(表 2,表 3);质谱条件:扫描方式多反应监测(multi-reaction monitoring, MRM,表 4),离子源温度150 ℃,去溶剂雾化流量800 L/h,雾化锥体流量150 L/h,雾化器压力7.0 bar,去溶剂温度350 ℃,毛细管喷涂电压3 kV。
| Condition | Glutinectin | Fengamycin | Natamycin |
| Mobile phase | 1.4 g/L trichloroacetic acid | Methanol | Methanol |
| Velocity (mL/min) | 1.0 | 0.3 | 0.6 |
| Temperature (℃) | 40.0 | 35.0 | 30.0 |
| Wavelength (nm) | 254.0 | 279.0 | 304.0 |
| Injection volume (μL) | 20.0 | 20.0 | 20.0 |
| Analyze time (min) | 20.0 | 20.0 | 20.0 |
| Time (min) | Velocity (mL/min) | Mobile phase A (%) | Mobile phase B (%) |
| 0.0 | 0.3 | 20.0 | 80.0 |
| 1.0 | 0.3 | 20.0 | 40.0 |
| 2.0 | 0.3 | 20.0 | 20.0 |
| 2.6 | 0.3 | 20.0 | 80.0 |
| 5.0 | 0.3 | 20.0 | 80.0 |
| t/min | Velocity (mL/min) | Mobile phase A (%) | Mobile phase B (%) |
| 0.0 | 0.3 | 90.0 | 10.0 |
| 2.0 | 0.3 | 90.0 | 10.0 |
| 6.0 | 0.3 | 10.0 | 90.0 |
| 8.0 | 0.3 | 10.0 | 90.0 |
| 8.5 | 0.3 | 90.0 | 10.0 |
| 10.0 | 0.3 | 90.0 | 10.0 |
| Substance | Ionic mode | Precursor ion (m/z) | Product ion (m/z) | Declustering potential (Ⅴ) | Collision energy (Ⅴ) |
| Glutinectin | ES+ | 465.0 | 455.8 | 152.9 | 27.3 |
| Fengamycin | ES+ | 292.1 | 160.3 | 108.0 | 24.0 |
| Natamycin | ES+ | 666.4 | 503.5 | 114.0 | 13.0 |
1.6 挥发性成分检测及鉴定
用顶空固相微萃取-气质联用的方法分析菌株K2的挥发性代谢产物。将在250 mL三角瓶中培养10 d的链霉菌麦粒培养基置于45 ℃恒温水浴30 min,插入经老化处理的DVB/CAR/ PDMS固相微萃取纤维头,顶空吸附30 min,使挥发性物质富集于萃取头上,然后直接进样到气相色谱-质谱(gas chromatograph-mass spectrometer, GC-MS)联用仪,于250 ℃下解析5 min后进行GC-MS分析,GC-MS操作条件参照文献[11]。计算机自动将所得气体成分质谱与国际标准数据库(NISTO17 Mass Spectrometry Library Database)数据进行比对,鉴定出挥性物质的成分,以测得的全部峰面积作为100%,用归一化法确定各组分的相对含量。以相同质量无菌麦粒培养基作为对照,将对照与菌株K2麦粒培养基中同时存在的物质去除,试验重复3次。
1.7 数据分析用MEGA7.0软件和邻接法构建系统发育树;用Microsoft Office Excel 2019和SPSS 20软件进行数据分析。
2 结果与分析 2.1 灭菌培养液的抗菌活性液体培养基成分对链霉菌次级代谢产物的产量有着重要的影响,为筛选更适合菌株K2的液体培养基,选择2种不同的培养基并采用管碟法比对培养液的抑菌活性,结果(表 5)表明,菌株K2在培养基A产生的次级代谢产物对病原真菌的抑菌活性均比培养基B好,更有利于代谢产物的产生。进一步与菌株CK-15进行抑菌活性比较,菌株K2对灰霉病菌的抑菌带略高于菌株CK-15,且菌株K2对苹果腐烂病菌、苹果轮纹病菌、核盘菌、杨树溃疡病菌、烟草赤星病菌的抑制作用强,抑菌带直径分别为20.7、15.7、15.3、14.3和14.0 mm,可见菌株K2的抑菌谱较广。
| Pathogens | K2 antagonistic band (mm) | CK-15 antagonistic band on culture media A (mm) | |
| Culture media A | Culture media B | ||
| Alternaria alternata | 14.0±1.0a | 12.7±0.6ab | 10.3±0.6b |
| Botryosphaeria dothidea | 14.3±0.6a | 13.3±0.6a | 14.3±1.2a |
| Botrytis cinerea | 13.5±1.5a | 12.5±0.7a | 11.0±1.7a |
| Exserohilum turcicum | 12.0±3.5a | 11.0±1.0a | 9.7±1.5b |
| Fusarium oxysporum | 5.7±0.0a | 5.0±0.6a | 5.0±1.0a |
| Physalospora piricola | 15.7±4.2a | 15.0±1.0a | 15.6±1.5a |
| Phytophthora capsici | 12.5±0.7a | 11.3±2.1a | 5.3±1.7b |
| Rhizoctonia cerealis | 8.3±2.1a | 7.6±0.6a | 4.0±1.2b |
| Rhodotorula rubra | 24.7±3.5a | 23.3±0.7a | 23.1±1.1a |
| Sclerotinia sclerotiorum | 15.3±2.3a | 13.3±0.6a | 13.3±1.0a |
| Valsa mali | 20.7±1.2a | 14.3±2.3b | 21.0±1.0a |
| Acidovorax citrulli | 0.0 | 0.0 | 0.0 |
| Pseudomonas syringae | 0.0 | 0.0 | 0.0 |
| Ralstonia solanacearum | 0.0 | 0.0 | 0.0 |
| Xanthomonas vesicatoria | 0.0 | 0.0 | 0.0 |
| Xanthomonas oryzae | 0.0 | 0.0 | 0.0 |
| Data were the mean of 3 replicates with standard deviation. Different lowercase letters in the same row indicate significant differences (P < 0.05), among different culture media tested with Duncan’s multiple range test. | |||
2.2 K2产挥发性物质对灰霉病菌的抑菌活性
25 ℃恒温培养箱中培养5 d后,对照组灰霉病菌几乎长满平板(图 1)。双皿对峙中,在菌株K2的挥发性物质作用下灰霉菌不生长,抑菌率为100% (表 6);在菌株CK-15的挥发性物质作用下灰霉菌落直径为11.0 mm,抑菌率为86.8%。用李其利[10]的方法测定,发现在相同挥发性物质的量下增大作用的面积后,挥发性物质对灰霉病菌的抑制效果减弱,菌株K2和CK-15处理后灰霉菌落直径分别为65.8、81.1 mm,抑菌率也降至仅20.1%、1.5%。由此可见,产生的挥发性物质对灰霉病菌生长的抑制效果与挥发性物质的量有关,并且与菌株CK-15相比菌株K2产的挥发性物质对灰霉病菌抑制效果更好。

|
| 图 1 挥发性物质对灰霉病菌菌丝生长的影响 Figure 1 Effect of volatile substances on mycelia growth of Botrytis cinerea. A–C: The result of the double-dish confrontation test. D–F: The results of the Li Qili's method. A, D: Sterile water control; B, E: Volatile substances produced by K2 on grain medium; C, F: Volatile substances produced by CK-15 on grain medium. |
| Strain | Double-dish confrontation method | Li Qili’s method | |||
| Colony diameter (mm) | Growth inhibition of hyphae (%) | Colony diameter (mm) | Growth inhibition of hyphae (%) | ||
| CK | 83.3±0.5a | 0.0 | 82.3±0.5a | 0.0 | |
| K2 | 0.0c | 100 | 65.8±2.5b | 20.1 | |
| CK-15 | 11.0±4.3b | 86.8 | 81.1±3.4a | 1.5 | |
| Different lowercase letters in the same column indicate significant differences (P < 0.05). | |||||
2.3 菌株的鉴定
通过PCR扩增获得了菌株K2的16S rRNA基因片段,其序列长度为1 465 bp。将该序列提交至NCBI数据库进行BLAST分析比对,结果(图 2)显示菌株K2与Streptomyces lydicus的相似性达到99%。在用MEGA 7.0构建的邻接系统发育树上,K2菌株与Streptomyces lydicus处于同一末端分支上(bootstrap支持率为99%),因此,将菌株K2鉴定为利迪链霉菌(Streptomyces lydicus)。

|
| 图 2 通过邻接法构建的基于K2菌株和相关链霉菌的16S rRNA基因序列的系统发育树 Figure 2 Phylogenic tree constructed with the neighbor-joining method based on 16S rRNA gene sequences of strain K2 and the related Streptomyces species. The tree was constructed with MEGA 7.0 software. The number at each node indicates the bootstrap support and the bar indicates expected number of substitutions per site. |
2.4 K2的培养液中活性成分定性分析
用高效液相色谱对菌株培养液进行定性检测分析的结果(图 3)表明,在HPLC检测结果中谷氏菌素、丰加霉素和纳他霉素标准品出峰保留时间分别为6.736、3.654、4.466 min,比对菌株K2培养液在相同检测条件下与谷氏菌素和丰加霉素标准品具有相同保留时间的峰,比纳他霉素标准品出峰保留时间滞后0.531 s。在LC-MS检测分析(图 4)的结果中谷氏菌素、丰加霉素和纳他霉素标准品出峰保留时间分别为7.36、1.59 min和3.73 min,比对菌株K2培养液在相同检测条件下具有相同保留时间的峰。因此证实菌株K2培养液次级代谢产物中含有谷氏菌素、丰加霉素和纳他霉素。

|
| 图 3 K2菌株培养液HPLC分析 Figure 3 HPLC analysis of culture broth from strain K2. A, B and C are the chromatographic diagrams of the standards of glutamicin, fongamycin and natamycin, respectively. D, E and F are the chromatograms of glutamicin, fongamycin and natamycin in the culture broth of strain K2 under the same detection conditions. |

|
| 图 4 K2菌株培养液LC-MS分析 Figure 4 LC-MS analysis of culture broth from strain K2. A and B are chromatograms of the standards of glutamicin and the culture broth of strain K2 under the same detection conditions, respectively; C and D are chromatographic diagrams of the standards of fongamycin and the culture broth of strain K2 under the same detection conditions, respectively; E and F are chromatographic diagrams of the standards of natamycin and the culture broth of strain K2 under the same detection conditions, respectively. |
2.5 挥发性成分检测及鉴定
用顶空固相微萃取-气质联用的方法检测后将检测结果与数据库进行比对,鉴定菌株K2挥发性代谢产物。结果(表 7)表明,菌株K2可产生30种挥发性物质,它们多数属于烯类、醇类、酯类和烷烃类,其中含量最高的是2-methylisoborneol、1-undecene和p-menth-8-ene,3-methylene的含量次之。在所获得物质中有已报道能够抑制菌核病菌、尖镰孢和水稻纹枯病菌菌丝生长的benzothiazole[12],还有对意大利青霉、白色念珠菌、大肠杆菌等有较强的抑菌活性的β-pinene[13]。所以,推测菌株K2产挥发性物质的抑菌活性可能与这几种物质有关。
| Compounds | Peak area percentage (%) |
| 2-pentanone | 0.10 |
| Silanediol, dimethyl- | 0.23 |
| 2-pentanol | 0.19 |
| 2-octynoic acid | 0.10 |
| β-pinene | 0.18 |
| 1,3-cyclopentadiene, 1,3-bis(1-methylethyl)- | 3.13 |
| 2,4-cycloheptadien-1-one, 2,6,6-trimethyl- | 1.38 |
| p-menth-8-ene, 3-methylene- | 15.82 |
| Benzene, 1-methoxy-4-methyl- | 0.83 |
| 1-pentene, 5-(2,2-dimethylcyclopropyl)-2-methyl-4-methylene- | 0.20 |
| 1,4-undecadiene, (E)- | 0.12 |
| 1-undecene | 8.23 |
| Benzene, 1,2-dimethoxy- | 0.18 |
| Benzene, 1,4-dimethoxy- | 1.05 |
| Benzene, 1,3-dimethoxy- | 0.10 |
| 2-methylisoborneol | 62.89 |
| Pentanoic acid, 1,3,3-trimethylbicyc lo[2.2.1]hept-2-yl ester | 0.27 |
| Oxirane, hexadecyl- | 0.10 |
| 1,7,7-trimethylbicyclo[2.2.1]hept-5-en-2-ol | 0.17 |
| Benzothiazole | 0.19 |
| 1,4-methano-1H-cyclohepta[d]pyridazine, 4,4a,5,6,7,8,9,9a-octahydro-10,10-dimethyl- | 1.18 |
| 1,4-methano-1H-cyclohepta[d]pyridazine, 4,4a,5,6,7,8,9,9a-octahydro-10,10-dimethyl- | 0.72 |
| Spiro[4.5]decane, 6-methylene- | 0.17 |
| 1,2,4-trimethoxybenzene | 0.24 |
| trans-1,10-dimethyl-trans-9-decalinol | 1.20 |
| Bicyclo[5.3.0]decane, 2-methylene-5-(1-methylvinyl)-8-methyl- | 0.16 |
| Thunbergol | 0.11 |
| Germacrene D | 0.43 |
| gamma.-elemene | 0.10 |
| 3,7-cyclodecadiene-1-methanol,a,a,4,8-tetramethyl-, [S-(E,E)]- | 0.23 |
3 讨论与结论
灰葡萄孢(B. cinerea)可侵染200多种植物,被列为第二重要的植物病原菌[14]。研究发现在灰霉病控制中长期单独使用某一类化学药剂易使灰葡萄孢产生抗药性,造成防治效果降低[15]。在绿色发展的倡导下,开发利用有益微生物进行生物防治是未来农药的发展的重要方向之一。放线菌是人类最早研究并利用的拮抗微生物,链霉菌作为放线菌的主要类群,在其生长发育过程中可产生多种对植物病原菌有抑制作用的次生代谢产物,在控制植物真菌和细菌病害方面具有相当大的潜力[16]。本研究对利迪链霉菌K2从培养液和产挥发性物质2种代谢途径进行抑菌活性探究,进一步分析了有效抑菌活性成分,表明K2可产生具有很强抑菌活性的挥发性物质,具有开发成生物农药和生物熏蒸剂的潜能,也为后续对菌株K2的深入研究奠定了基础。
目前,在生物防治方面,利迪链霉菌主要作为植物根际促生菌,对西瓜、豌豆、棉花和玉米等作物均有较好的促生效果,且对镰刀菌(Fusarium spp.)、腐霉菌(Phytium spp.)和丝核菌(Rhizoctonia sp.)等病原真菌具有良好抑制作用[17-18]。Deobald和Sabaratnam等[19-20]报道了利迪链霉菌的孢子制成的菌剂可以用来防治豌豆根腐病(P. ultimum)、番茄猝倒病(R. solani)等多种土传病害。而目前关于利迪链霉菌防治灰霉病的研究报道还不多。谢晨昭等[21]利用淡紫灰链霉菌B1防治番茄灰霉病,其发酵上清液在离体叶片和活体植株上的防效分别为68.8%和64.0%;单丽萍等[22]的研究发现白黄链霉TD-1发酵液可以抑制灰霉病菌的孢子萌发,提高番茄多酚氧化酶、过氧化物酶的酶活力,从而促进番茄更好地抵御植物病原真菌;唐德链霉菌发酵液对草莓灰霉病菌、桃褐腐病菌和黄瓜枯萎病菌等植物病原菌具有明显的抑菌活性[23];链霉菌LA-5培养液对番茄果实灰霉病的防效可达83.4%[24]。生防链霉菌的研究主要集中在其次生代谢过程中所产生的对病原菌有抑制作用的抗生素的筛选与利用,应用于防治植物病害的抗生素有春雷霉素、链霉素、武夷菌素、中生菌素和多抗霉素等[25]。本研究中发现K2产生的次级代谢产物对灰霉病菌的抑制效果略高于菌株CK-15,对苹果腐烂病菌、苹果轮纹病菌、核盘菌、杨树溃疡病菌和烟草赤星病菌均具有较强的拮抗作用。高效液相色谱及液相色谱-质谱定性检测到该菌培养液中含有谷氏菌素、丰加霉素和纳他霉素。因此利迪链霉菌K2的次级代谢产物具有一定的开发潜能。
文献中已报道了多种链霉菌产生的VOCs,它们能显著抑制病原菌的生长从而有效的防治植物病害。邢梦玉[26]从土壤中筛选出链霉菌菌株TJGA-19、BWL-H1,产生的VOCs不仅能抑制荔枝霜疫霉菌丝生长,还能有效地预防荔枝霜疫病的发生,且BWL-H1产生的VOCs抑菌谱较广,能抑制15种病原菌;李其利[10]的研究证实链霉菌JK-1产生的VOCs能够导致灰霉菌分生孢子异常,并能有效控制储藏期番茄的灰霉病和柑橘的青霉病;万明国[27]发现普特拉链霉菌F-1产生的VOCs对水稻纹枯病、油菜菌核病和草莓灰霉病以及由细菌引起的青椒储藏期病害具有明显的防治效果;吕昂[28]发现杨氏链霉菌3-10菌株小麦粒培养物用量为17 g/L时,能显著抑制黄曲霉和寄生曲霉侵染花生,用量为86 g/L时,几乎能完全抑制这2种真菌的生长繁殖,花生粒上也检测不到黄曲霉毒素。本研究证实了利迪链霉菌K2在麦粒培养基上产挥发性物质对灰霉病菌有较好的抑制效果,且这种抑菌作用强弱与K2产挥发性物质的量有关,挥发性成分中具有benzothiazole和β-pinene,这为用利迪链霉菌K2研制果蔬贮藏期病害的生物熏蒸剂提供参考依据。
综上所述,利迪链霉菌K2具有良好的研发和应用潜力,是一株能有效防治灰霉病害的多功能菌种,可以用其开发成生产上防治灰霉病害的生防菌剂和防治采后储藏期果蔬灰霉病害的生物熏蒸剂。本研究进行的菌株抑菌活性测定为用K2研制新生物农药提供了依据,但今后对有关菌株发酵工艺、毒理试验和田间防效、菌株K2产挥发性物质的应用及其作用机理等仍需进一步深入试验研究。
| [1] |
TONG YH, JI ZL, XU JY, CHEN XJ. Research progress on biological control of gray mold. Chinese Journal of Biological Control, 2003(3): 131-135.
(in Chinese) 童蕴慧, 纪兆林, 徐敬友, 陈夕军. 灰霉病生物防治研究进展. 中国生物防治, 2003(3): 131-135. |
| [2] |
ZHANG L, ZHANG BY, HUANG JH, LI YG, YANG YJ. Research progress on biological control of Botrytis cinerea. Modern Animal Husbandry, 2021, 5(2): 55-60.
(in Chinese) 张莉, 张博源, 黄继红, 李琰歌, 杨艳君. 植物灰霉病生物防治研究进展. 现代牧业, 2021, 5(2): 55-60. |
| [3] | HOU J, GAO YN, FENG J, MA Z, ZHANG X. Sensitivity of Botrytis cinerea to propamidine: in vitro determination of baseline sensitivity and the risk of resistance. European Journal of Plant Pathology, 2010, 128(2): 261-267. DOI:10.1007/s10658-010-9661-6 |
| [4] |
WANG WL, JIN H, CONG BC, ZHOU L, WEI Z, WANG SM. Biocontrol effect of composite microbial agent on tomato bacterial wilt. Journal of Nanjing Agricultural University, 2022, 45(6): 1174-1182.
(in Chinese) 王文丽, 金涵, 从炳成, 周蕾, 韦中, 王世梅. 复合微生物菌剂对番茄青枯病的生防效应. 南京农业大学学报, 2022, 45(6): 1174-1182. |
| [5] |
WU Z, SUN L, LIU YY, WANG JW, ZHANG KC. Effects of bio-pesticide wuyiencin on controlling tomato gray mold in protected field. Chinese Agricultural Science Bulletin, 2013, 29(25): 173-178.
(in Chinese) 武哲, 孙蕾, 刘彦彦, 王家旺, 张克诚. 生物农药武夷菌素对保护地番茄灰霉病的防治效果. 中国农学通报, 2013, 29(25): 173-178. |
| [6] | CAZORLA FM, ROMERO D, PÉREZ-GARCÍA A, LUGTENBERG B, VICENTE AD, BLOEMBERG GV. Isolation and characterization of antagonistic Bacillus subtilis strains from the avocado rhizoplane displaying biocontrol activity. Journal of Applied Microbiology, 2010, 103(5): 1950-1959. |
| [7] | TYC O, SONG C, DICKSCHAT JS, VOS M, GARBEVA P. The ecological role of volatile and soluble secondary metabolites produced by soil bacteria. Trends in Microbiology, 2017, 25(4): 280-292. DOI:10.1016/j.tim.2016.12.002 |
| [8] |
SAPERS GM, GORNY JR, YOUSEF AE. Microbiology of Fruits and Vegetables[M]. CHEN W, TIAN FW, trans. Beijing: China Light Industry Press, 2011 (in Chinese). SAPERS GM, GORNY JR, YOUSEF AE. 果蔬微生物学[M]. 陈卫, 田丰伟译. 北京: 中国轻工业出版社, 2011. |
| [9] |
MA XR, SU DM. Microbiological Assay of Antibiotics. Beijing: Science Press, 2001. (in Chinese) 马绪荣, 苏德模. 抗生素微生物检定法. 北京: 科学出版社, 2001. |
| [10] |
LI QL. Identification, biocontrol potential and mode of action of Streptomyces JK-1[D]. Wuhan: Master's Thesis of Huazhong Agricultural University, 2011 (in Chinese). 李其利. 链霉菌JK-1的鉴定及其防病潜能和防病机制的研究[D]. 武汉: 华中农业大学博士学位论文, 2011. |
| [11] |
ZHANG CH. Study on the inhibiting of volatiles produced by Streptomyces alboflavus TD-1 on Botrytis cinerea[D]. Tianjin: Master's Thesis of Tianjin University of Science & Technology, 2015 (in Chinese). 张春慧. 链霉菌TD-1产挥发性物质抑制番茄灰霉病菌的研究[D]. 天津: 天津科技大学硕士学位论文, 2015. |
| [12] | RAZA W, YUAN J, LING N, HUANG Q, SHEN Q. Production of volatile organic compounds by an antagonistic strain Paenibacillus polymyxa WR-2 in the presence of root exudates and organic fertilizer and their antifungal activity against Fusarium oxysporum f. sp. niveum. Biological Control, 2015, 80: 89-95. DOI:10.1016/j.biocontrol.2014.09.004 |
| [13] |
ZHANG MH, WANG M, YANG SZ, SU XJ, DAI M, PENG LT. Possible action mode of beta-pinene against Penicillium italicum. Journal of Huazhong Agricultural University, 2018, 37(6): 91-97.
(in Chinese) 张美红, 王萌, 杨书珍, 苏小军, 代梅, 彭丽桃. β-蒎烯抑制柑橘意大利青霉作用机制初步研究. 华中农业大学学报, 2018, 37(6): 91-97. |
| [14] | R DEAN, JALV KAN, ZA PRETORIUS, KE HAMMOND-KOSACK, AD PIETRO, PD SPANU, JJ RUDD, M DICKMAN, R KAHMANN, J ELLIS. The top 10 fungal pathogens in molecular plant pathology. Molecular Plant Pathology, 2012, 13(4): 414-430. |
| [15] |
ZHANG W, QIAO GH, HUANG JB, WANG ZY, LI XH. Evaluation on resistance of grape gray mold pathogen Botrytis cinerea to pyrimethanil in China. Scientia Agricultura Sinica, 2013, 46(6): 1208-1212.
(in Chinese) 张玮, 乔广行, 黄金宝, 王忠跃, 李兴红. 中国葡萄灰霉病菌对嘧霉胺的抗药性检测. 中国农业科学, 2013, 46(6): 1208-1212. |
| [16] |
DAI Y, WU XR, CHEN YJ. Advances in strategies for activating silent biosynthetic gene clusters in Streptomyces. Journal of China Pharmaceutical University, 2019, 50(4): 379-388.
(in Chinese) 戴岩, 吴旭日, 陈依军. 链霉菌沉默生物合成基因簇激活策略的研究进展. 中国药科大学学报, 2019, 50(4): 379-388. |
| [17] | YUAN WM, CRAWFORD DL. Characterization of Streptomyces lydicus WYEC 108 as a potential biocontrol agent against fungal root and seed rots. Applied and Environmental Microbiology, 1995, 61(8): 3119-3128. |
| [18] |
SUN HB, LI MY, WU HL, GUO SG, ZHANG J, REN Y, ZHANG HY, GONG GY, XU Y. Biocontrol and plant growth promoting effects of Streptomyces against Fusarium wilt in watermelon. Science Technology and Engineering, 2020, 20(13): 5074-5079.
(in Chinese) 孙洪宝, 李茂营, 吴慧玲, 郭绍贵, 张洁, 任毅, 张海英, 宫国义, 许勇. 生防菌链霉菌对西瓜枯萎病防治及幼苗生长的影响. 科学技术与工程, 2020, 20(13): 5074-5079. |
| [19] | DEOBALD LA. Isolation and characterization of Streptomyces strains antagonistic to phytopathogenic fungi and analysis of antibiotics potentially involved in disease control[D]. America: Doctoral Dissertation of University of Idaho, 2004. |
| [20] | SABARATNAM S, TRAQUAIR JA. Formulation of a Streptomyces biocontrol agent for the suppression of Rhizoctonia damping-off in tomato transplants. Biological Control, 2002, 23(3): 245-253. |
| [21] |
XIE CZ, YANG YL, LI L, LI JY, WANG JH, WANG HM. Taxonomy of antagonistic strain B1 and control effect of tomato gray mould. Acta Phytophylacica Sinica, 2008(4): 300-306.
(in Chinese) 谢晨昭, 杨毅玲, 李磊, 李金云, 王建辉, 王慧敏. 拮抗放线菌B1菌株鉴定及其防治番茄灰霉病的初步研究. 植物保护学报, 2008(4): 300-306. |
| [22] |
SHAN LP, WANG CL, LI ZJ, CHEN MH, LI FJ. Inhibitory effect of Streptomyces alboflavus TD-1 against Botrytis cinerea and its effect on defensive enzymes activity. Acta Agriculture Boreali-Sinica, 2015, 30(2): 100-103.
(in Chinese) 单丽萍, 王昌禄, 李贞景, 陈勉华, 李风娟. 链霉菌TD-1对番茄灰霉病菌的抑制及防御酶活性的影响. 华北农学报, 2015, 30(2): 100-103. |
| [23] |
ZHAO J, JIA WG, LIU WC, LIU DW, ZHANG DP, LU CG. Screening of antagonistic actinomycete against strawberry grey mold and activity determination of its fermentation broth. Northern Horticulture, 2018(4): 59-65.
(in Chinese) 赵娟, 贾卫国, 刘伟成, 刘德文, 张殿朋, 卢彩鸽. 草莓灰霉病菌拮抗放线菌的筛选及活性测定. 北方园艺, 2018(4): 59-65. |
| [24] |
LI PQ, FENG BZ, LI XX, HAO HY. Screening and identification of antagonistic actinomycete LA-5 against Botrytis cinerea. Chinese Journal of Applied Ecology, 2018, 29(12): 4172-4180.
(in Chinese) 李培谦, 冯宝珍, 李新秀, 郝号永. 番茄灰霉菌拮抗放线菌LA-5的筛选及鉴定. 应用生态学报, 2018, 29(12): 4172-4180. |
| [25] | IKENO S, AOKI D, HAMADA M, HORI M, TSUCHIYA KS. DNA sequencing and transcriptional analysis of the kasugamycin biosynthetic gene cluster from Streptomyces kasugaensis M338-M1. The Journal of Antibiotics, 2006, 59(1): 18-28. |
| [26] |
XING MY. Study on biocontrol potential and antifungal mechanism of Streptomyces TJGA-19 and BWL-H1 against litchi downy blight[D]. Guangzhou: Master's Thesis of South China Agricultural University, 2017 (in Chinese). 邢梦玉. 两株链霉菌对荔枝霜疫病的防病潜力和防病机理研究[D]. 广州: 华南农业大学博士学位论文, 2017. |
| [27] |
WANG MG. Characterization of the identity and biocontrol potential of astrain of Streptomyces F-1isolated from rice[D]. Wuhan: Master's Thesis of Huazhong Agricultural University, 2008 (in Chinese). 万明国. 源于水稻的链霉菌菌株F-1的鉴定及其防病潜力和防病机制研究[D]. 武汉: 华中农业大学博士学位论文, 2008. |
| [28] |
LV A. Identification of the antifungal metabolites from Streptomyces sp. 3-10 and evaluation of their efficacy against plant fungal diseases[D]. Wuhan: Master's Thesis of Huazhong Agricultural University, 2017 (in Chinese). 吕昂. 链霉菌3-10抗真菌代谢产物鉴定及防病潜力评估[D]. 武汉: 华中农业大学博士学位论文, 2017. |