 2023, Vol. 50
2023, Vol. 50
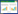



阻断消耗途径提高毕赤酵母工程菌S-腺苷甲硫氨酸产量

扩展功能
文章信息
- 苏慧颖, 蒙世拯, 赵欣欣, 钱江潮, 储炬, 秦秀林
- SU Huiying, MENG Shizheng, ZHAO Xinxin, QIAN Jiangchao, CHU Ju, QIN Xiulin
- 阻断消耗途径提高毕赤酵母工程菌S-腺苷甲硫氨酸产量
- Blocking consumption pathway increases production of S-adenosyl-L-methionine by Pichia pastoris
- 微生物学通报, 2023, 50(2): 441-453
- Microbiology China, 2023, 50(2): 441-453
- DOI: 10.13344/j.microbiol.china.220486
-
文章历史
- 收稿日期: 2022-05-15
- 接受日期: 2022-06-30
- 网络首发日期: 2022-07-21
苏慧颖, 蒙世拯, 赵欣欣, 钱江潮, 储炬, 秦秀林. 阻断消耗途径提高毕赤酵母工程菌S-腺苷甲硫氨酸产量[J]. 微生物学通报, 2023, 50(2): 441-453.

SU Huiying, MENG Shizheng, ZHAO Xinxin, QIAN Jiangchao, CHU Ju, QIN Xiulin. Blocking consumption pathway increases production of S-adenosyl-L-methionine by Pichia pastoris[J]. Microbiology China, 2023, 50(2): 441-453.
阻断消耗途径提高毕赤酵母工程菌S-腺苷甲硫氨酸产量
1. 广西大学生命科学与技术学院 亚热带农业生物资源保护与利用国家重点实验室 广西微生物与酶工程技术研究中心, 广西 南宁 5300042;
2. 华东理工大学 生物反应器工程国家重点实验室, 上海 200237
2. 华东理工大学 生物反应器工程国家重点实验室, 上海 200237
摘要: 【背景】 S-腺苷甲硫氨酸(S-adenosyl-L-methionine, SAM)作为所有生物体内的重要中间代谢物,不仅可作为膳食补充剂,还具有良好的临床应用价值。【目的】 将毕赤酵母重组菌GS115/DS16的SAM消耗途径阻断,进一步提高SAM的产量。【方法】 分别敲除毕赤酵母重组菌GS115/DS16的S-腺苷同型半胱氨酸水解酶基因sah1、S-腺苷甲硫氨酸脱羧酶基因spe2和L-甲硫氨酰tRNA合酶基因msm1,构建工程菌G/Dsah、G/Dspe和G/Dmsm。检测3个工程菌的生长和SAM产量,以及l-Met添加量对SAM积累的影响。【结果】 与出发菌GS115/DS16相比,工程菌G/Dsah、G/Dspe和G/Dmsm的单位菌体SAM产量分别提高了29.3%、55.6%和24.8%,其生长无显著差异。l-Met添加量优化后(0.06%),G/Dsah和G/Dmsm单位菌体的SAM产量分别提高了26.4%和28.9%。【结论】 构建的毕赤酵母工程菌可用于SAM的工业化生产,该代谢工程策略可用于改进其他化学品的生产。
关键词:
S-腺苷甲硫氨酸 毕赤酵母 S-腺苷甲硫氨酸脱羧酶 S-腺苷同型半胱氨酸水解酶 L-甲硫氨酰tRNA合酶
Blocking consumption pathway increases production of S-adenosyl-L-methionine by Pichia pastoris
1. State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources, Guangxi Research Center for Microbial and Enzyme Engineering Technology, College of Life Science and Technology, Guangxi University, Nanning 530004, Guangxi, China;
2. State Key Laboratory of Bioreactor Engineering, East China University of Science and Technology, Shanghai 200237, China
2. State Key Laboratory of Bioreactor Engineering, East China University of Science and Technology, Shanghai 200237, China
Abstract: [Background] S-Adenosyl-L-methionine (SAM) is an important intracellular metabolite that can be used as a dietary supplement and to treat a variety of diseases. [Objective] To enhance SAM production for industrial application by blocking SAM consumption pathways in the recombinant Pichia pastoris strain GS115/DS16. [Methods] The genes associated with the metabolism of SAM, sah1 (encoding S-adenosyl-L-homocysteine hydrolase), spe2 (encoding S-adenosylmethionine decarboxylase), and msm1 (encoding mitochondrial methylthio-tRNA synthase), were knocked out in SAM-producing strain GS115/DS16. Accordingly, the engineered strains G/Dsah, G/Dspe, and G/Dmsm were constructed. The cell growth and SAM production of the three engineered strains were investigated. Additionally, the effect of methionine addition on SAM accumulation was studied. [Results] The knock-out did not affect cell growth, whereas it increased SAM production by 29.3%, 55.6%, and 24.8% in G/Dsah, G/Dspe, and G/Dmsm, respectively, compared with the parental strain GS115/DS16. When l-Met addition was decreased from 0.10% to 0.06%, the SAM production increased by 26.4% and 28.9% in G/Dsah and G/Dmsm, respectively. [Conclusion] Therefore, the engineered P. pastoris strains can be utilized in industrial production of SAM in a cost-effective manner, and the strategy can also be employed for improving the production of other chemicals.
Keywords:
S-adenosyl-L-methionine Pichia pastoris S-adenosylmethionine decarboxylase S-adenosyl-L-homocysteine hydrolase mitochondrial methionyl-tRNA synthetase
S-腺苷甲硫氨酸(S-adenosyl-L-methionine, SAM)作为所有生物体内的重要中间代谢物,是一种高价值的化学物质,具有良好的临床应用价值。SAM不仅被广泛地用作膳食补充剂,也是抑郁症、肝病和癌症的有效治疗剂[1-2]。利用代谢工程改造微生物细胞的代谢途径可显著提高SAM在细胞内的积累[3-6]。主要的改造方法有:(1) 强化S-腺苷甲硫氨酸合成酶(methionine adenosyltransferase, MAT)基因(sam1和sam2)的表达量,提高MAT的活性,从而增加SAM的产量;(2) 阻断或弱化β-胱硫醚合成酶基因(cys4)表达,减少SAM和L-Met在细胞内转化为半胱氨酸的途径,增加胞内SAM的积累;(3) 减少SAM分解代谢(消耗)途径,提高SAM的胞内积累;(4) 增加ATP、L-甲硫氨酸(L-Met)或前体的量,以提高SAM的胞内积累。
在生物体内MAT催化ATP和L-Met合成SAM,不同来源的MAT基因的过表达是提高SAM产量的首要策略[7]。当MAT活性提高到一定水平时,它不再是SAM合成的限速因子[8]。因此,在获得了MAT活性足够高的工程菌后,减少SAM分解代谢途径是增加细胞内SAM积累的另一有效策略。
在毕赤酵母(Pichia pastoris)中,SAM代谢途径如图 1A所示。L-Met和ATP在MAT的作用下反应生成SAM,SAM可通过S-腺苷甲硫氨酸脱羧酶(S-adenosylmethionine decarboxylase, SPE2)催化的脱羧反应生成脱羧S-腺苷甲硫氨酸;也可在S-腺苷高半胱氨酸水解酶(S-adenosyl-L-homocysteine hydrolase, SAH1)作用下催化S-腺苷高半胱氨酸(S-adenosyl-L-homocysteine, AdoHcy)形成高半胱氨酸,随后转化为β-胱硫醚或L-Met[9]。L-Met作为参与SAM合成的底物,在L-甲硫氨酰tRNA合酶(mitochondrial methionyl-tRNA synthetase, MSM1)的作用下[10],先形成L-甲硫氨酰-tRNA,再反应生成N-甲酰-甲硫氨酰-tRNA,参与生理反应。我们前期的研究发现,弱化S-腺苷甲硫氨酸(S-adenosyl-L-methionine, SAM)分解代谢途径β-胱硫醚(β-cystathionine)合成途径能提高毕赤酵母重组菌的SAM产量[4],因此,减少SAM在细胞内的消耗是提高其产量的有效策略。

|
| 图 1 毕赤酵母重组工程菌G/Dspe、G/Dsah和G/Dmsm的构建 Figure 1 Construction of recombinant strains G/Dspe, G/Dsah and G/Dmsm. A:毕赤酵母SAM代谢途径. sah1:S-腺苷同型半胱氨酸水解酶基因;spe2:S-腺苷甲硫氨酸脱羧酶基因;sam:SAM合成酶基因;msm1:L-甲硫氨酰tRNA合酶基因;cys4:β-胱硫醚合成酶基因. B:重组质粒pDspeZ、pDsahZ和pDmsmZ的酶切验证. C:菌落PCR验证工程菌G/Dspe、G/Dsah和G/Dmsm A: Intracellular metabolic pathway for SAM utilization. The pathways of SAM metabolism in Pichia pastoris. sah1: S-adenosyl-L-homocysteine hydrolase gene; spe2: S-adenosylmethionine decarboxylase gene; sam: S-adenosylmethionine synthetase gene; msm1: Mitochondrial methionyl-tRNA synthetase gene; cys4: Cystathionine β-synthase gene. B: Verification of plasmids pDspeZ, pDsahZ and pDmsmZ by restriction enzyme digestion. C: Verification of engineered strains G/Dspe, G/Dsah and G/Dmsm by colony PCR. |
|
|
L-Met和ATP都是合成SAM的底物,增加底物浓度或降低其成本对SAM的合成有着重要的意义。构建以腺苷为底物合成ATP的重组菌可提高SAM产量并降低其生产成本[11-12]。L-Met既是合成SAM的底物,也是菌体生长的碳源和氮源,因此,L-Met的添加不仅影响SAM产量,还会对菌体的细胞代谢产生影响。L-Met作为SAM合成过程中需外源添加的底物,通常采用过量添加的方法,这种方法就会导致成本上升。有研究表明,胞内浓度过高的L-Met会抑制TCA循环和氧化磷酸化作用,造成SAM产量降低[13]。通过优化L-Met的补料策略[14],在突变酿酒酵母中使用低成本的dL-Met (混合的d-Met和L-Met)作为SAM生产的底物[15],能显著提高SAM产量。酵母细胞中L-Met的转运主要是利用MUP1和MUP3编码的甲硫氨酸透过酶。在毕赤酵母重组菌中,共表达腺苷酸激酶基因ADK1、MUP1和SAM2能提高L-Met转化效率和SAM的产量[16]。在酵母菌株中,共表达编码甲硫氨酸合成酶的Met6和SAM2可提高细胞内的L-Met水平,从而提高L-Met的利用率[17],降低SAM生产的成本。
S-腺苷同型半胱氨酸水解酶(SAH1)、S-腺苷甲硫氨酸脱羧酶(SPE2)和L-甲硫氨酰tRNA合酶(MSM1)是SAM代谢途径的关键酶,阻断这些途径可以减少SAM在胞内的消耗,提高工程菌积累SAM的能力。细胞内适度的L-Met水平有利于SAM的合成,优化L-Met添加浓度,可以提高工程菌SAM的产量。
1 材料与方法 1.1 菌株和质粒毕赤酵母(Pichia pastoris)表达质粒pGAPZ A和菌株GS115购自Invitrogen公司。大肠杆菌(Escherichia coli) DH5α保藏于本实验室,用于基因克隆,本研究所构建的其他质粒和菌株见表 1。
表 1 本研究所构建质粒和菌株
Table 1 Plasmids and strains used in this study
| Plasmids or strains | Relevant features | Source or reference |
| Plasmids | ||
| pGAPZ A | P. pastoris expression plasmid, PGAP promoter; ble+ | Invitrogen |
| pDspeZ | The spe2 gene knock out vector containing 5’spe2-ble-3’spe2 cassette; ble+ | This study |
| pDsahZ | The sah1 gene knock out vector containing 5’sah1-ble-3’sah1 cassette; ble+ | This study |
| pDmsmZ | The msm1 gene knock out vector containing 5’msm1-ble-3’msm1 cassette; ble+ | This study |
| Strains | ||
| GS115 | Commercial P. pastoris host strain; his4– | Invitrogen |
| GS115/DS16 | GS115 harboring recombinant methionine adenosyltransferase gene DS16; HIS4; kan+ | [18] |
| G/Dspe | The spe2 gene was knock out in GS115/DS16; spe2; ble+ | This study |
| G/Dsah | The sah1 gene was knock out in GS115/DS16; sah1; ble+ | This study |
| G/Dmsm | The msm1 gene was knock out in GS115/DS16; msm1; ble+ | This study |
培养基YPG、YPD、MD、BMGY、BMMY按照毕赤酵母表达系统说明书制备。
LLB培养基(g/L):胰蛋白胨10.0,酵母提取物5.0,NaCl 10.0,Zeocin 50 μg/mL。
YPDS培养基(g/L):胰蛋白胨20.0,酵母提取物10.0,葡萄糖20.0,山梨醇182.2,琼脂15.0。
BSM发酵培养基(g/L):K2SO4 18.20,CaSO4 0.93,85% H3PO4 26.80,MgSO4·7H2O 14.90,KOH 4.13,glycerol 40.00,毕赤微量元素溶液(Pichia trace minerals 1 salt solution, PTM1) 12 mL。
PTM1 (g/L):ZnCl2 20.00,KI 0.08,CoCl2 0.50,CuSO4·5H2O 6.00,MnSO4·H2O 3.00,Na2MoO4·2H2O 0.20,H3BO3 0.20,FeSO4·7H2O 65.00,生物素0.20,H2SO4 5.00。
1.3 主要试剂和仪器酵母氮源基础(yeast nitrogen base, YNB)、生物素、氨苄青霉素(ampicillin, Amp)、遗传霉素G418,Solarbio公司;ZeocinTM,Invitrogen公司;质粒小规模提取试剂盒、通用型DNA纯化回收试剂盒、RNA提取试剂盒,TIANGEN公司;2×Phanta® Max Master Mix、2×Taq Plus Master Mix、Exnase Ⅱ工具酶,Vazyme公司;限制性内切酶和DNA Marker,Thermo Fisher Scientific公司;T4 DNA Ligase,TaKaRa公司。气相色谱仪,上海海欣色谱仪器有限公司。
1.4 引物研究所用引物见表 2,由生工生物工程(上海)股份有限公司合成。
表 2 本研究所用引物
Table 2 Primers used in this study
| Primer name | Sequence (5′→3′) | Amplified fragment length (bp) |
| sah3-F | AGCGTCGACGGTTGAAGGCCAACGCTCAGGACGT | 490 |
| sah3-R | CGCGGATCCAAAGGAGGCAAAAGCTCTGTAGGTAATTAGC | |
| sah5-F | AGGACTAGTGAGACAAGTATACTGATCTTCTTTCTACCTATT | 970 |
| sah5-R | AAGGCGGCCGCGGTATTATTTGGTTTAGTGTAGTAGAGAGG | |
| msm3-F | AACGTCGACCTGTGACTTCACAGAAAGGAAACTAAAC | 516 |
| msm3-R | CGCGGATCCGGCGTAGAGCCCTTGAGAAATTTT | |
| msm5-F | AGGACTAGTCAGCAGTTTGGTCAATACAACCCTG | 997 |
| msm5-R | AAGGCGGCCGCACGTAGATGGTAAATACAGTGGTTAAGTGC | |
| spe3-F | AACGAGCTCGCGTCGACTTGGAATGAAAACTTTGAAAGAGACGG | 513 |
| spe3-R | CGCGGATCCATGTTACGAGGCTGTACTGAAATATTTGG | |
| spe5-F | AGGACTAGTCGGGAAATGACATCCTATAGGCGAATC | 661 |
| spe5-R | AACGAGCTCGGCGGCCGCATTGCCGGGATTAGAATGAAAAATTGTG | |
| zeocin-F | AAAGCGGCCGCTAACCCACACACCATAGCTTCAAAATGT | 1 780 |
| zeocin-R | AACGTCGACAGCTTGCAAATTAAAGCCTTCGAG | |
| sah5-F0 | CTTGCCAATTTTTCAAAAGCAATTC | 1 106 |
| zeocin R0 | ACATTTTGAAGCTATGGTGTGTGGG | |
| msm5-F0 | GGGGAATGTCTTTCTATCAACAACC | 1 100 |
| zeocin R0 | ACATTTTGAAGCTATGGTGTGTGGG | |
| spe5-F0 | GGCCTTCCTATAATGCGATGACTTC | 861 |
| zeocin R0 | ACATTTTGAAGCTATGGTGTGTGGG |
挑取MD平板活化的毕赤酵母重组菌GS115/DS16单菌落接种至YPD培养基中,于30 ℃、220 r/min条件下培养过夜;以1%接种量转接至100 mL YPD培养基,培养至OD600约为0.8−1.2,用于感受态细胞制备。毕赤酵母感受态细胞的制备和电击转化参照毕赤酵母表达手册进行。质粒pDsahZ、pDspeZ、pDmsmZ分别经SpeⅠ/BamHⅠ酶切线性化,然后电击转化GS115/DS16感受态细胞。电转后迅速加入1 mL冰预冷的无菌1 mol山梨醇,温和混匀,取100 μL涂布于YPDS平板(含100 μg/mL Zeocin),于30 ℃培养3 d,筛选转化子。
1.6 毕赤酵母工程菌摇瓶发酵生产SAM从MD平板上挑取单菌落接种至3 mL YPG培养基,30 ℃、220 r/min培养过夜,取1 mL培养液于25 mL BMGY中进行种子扩大培养,培养16 h (OD600约为20.0)。收集BMGY培养的种子液,于4 000 r/min离心5 min,弃上清,菌体重悬于25 mL BSM培养基(250 mL摇瓶),于30 ℃、220 r/min甲醇诱导培养96 h。每隔12 h用气相色谱测定甲醇残留浓度,添加甲醇至终浓度为1.2% (体积分数);每隔24 h添加一定量的L-Met,用5 mol/L氢氧化钾调节pH (5.5−6.0)。各工程菌分别选取6个转化子用摇瓶发酵,设3次生物学重复。
1.7 SAM浓度和SAM合成酶酶活检测胞内SAM浓度和SAM合成酶(MAT)的酶活测定用HPLC检测[19],采用Thermo-BioBasic SCX色谱柱(4.6 mm×250 mm, 5 µm)。流动相为:A:5 mmol/L甲酸铵,pH 4.0;B:500 mmol/L甲酸铵,pH 4.0。洗脱方式:0.0−5.0 min,100% A;5.01−9.00 min,10% A,90% B;9.01−12.00 min,100% B;12.01−14.00 min,100% A。设定流速为1 mL/min,检测波长254 nm。1个MAT酶活单位定义为:37 ℃条件下,1 h内转化生成1 µmol的SAM所对应的酶量。
1.8 气相色谱测定甲醇浓度每隔12 h取BSM发酵培养液1 mL,常温下8 000 r/min离心2 min,取上清,经滤头(0.22 µm)过滤,用于甲醇浓度测定。使用气相色谱仪,填料为chromosorbl01型,色谱柱长l m、内径2 mm,柱炉温度125 ℃,汽化室温度170 ℃,检测器温度170 ℃,选用H2作为火焰检测器燃气(流量为30 mL/min),空气为助燃气(流量为300 mL/min),N2作为载气(流量为15 mL/min),采用CDMC色谱工作站进行分析。
2 结果与分析 2.1 敲除spe2、sah1和msm1的工程菌构建 2.1.1 基因敲除质粒的构建阻断SAM在胞内的消耗途径(图 1A)能进一步提高毕赤酵母工程菌GS115/DS16[18]的SAM产量。为了敲除DS16的S-腺苷同型半胱氨酸水解酶基因sah1、S-腺苷甲硫氨酸脱羧酶基因spe2和L-甲硫氨酰tRNA合成酶基因msm1,构建了基因敲除载体pDsahZ、pDspeZ和pDmsmZ。首先,以毕赤酵母菌GS115/DS16基因组DNA为模板,用引物对spe5-F/spe5-R、spe3-F/spe3-R分别扩增基因spe2 5′端和3′端片段。然后,将基因spe2 5′端和3′端片段分别克隆到质粒pGAPZ A,构建质粒pDspe。最后,以pGAPZ A质粒为模板,用引物对zeocinF/zeocinR扩增zeocin抗性基因ble片段,并克隆到pDspe质粒的NotⅠ/SalⅠ位点,获得用于敲除spe2的质粒pDspeZ。
以GS115/DS16的基因组DNA为模板,用引物对sah5-F/sah5-R、sah3-F/sah3-R分别扩增sah1的5′端和3′端片段;分别将sah1的5′和3′片段替换载体pDspeZ中spe2的5′和3′片段,获得质粒pDsahZ。同样地,分别将msm1的5′和3′片段替换载体pDspeZ中spe2的5′和3′片段,构建质粒pDmsmZ。质粒pDspeZ和pDsahZ、pDmsmZ的酶切验证正确(图 1B)。
2.1.2 基因敲除工程菌的构建质粒pDspeZ、pDsahZ和pDmsmZ经SpeⅠ/BamHⅠ酶切线性化,分别电击转化GS115/DS16,获得工程菌G/Dspe、G/Dsah和G/Dmsm。分别用引物对zeocin R0/sah5-F0、zeocin R0/msm5-F0和zeocin R0/spe5-F0对G/Dspe、G/Dsah和G/Dmsm转化子进行PCR验证。对应的工程菌G/Dsah和G/Dmsm扩增出1.1 kb片段,G/Dspe扩增出0.8 kb片段(图 1C)。
2.2 阻断SAM分解代谢途径对工程菌生长的影响在MD平板上,工程菌G/Dspe、G/Dmsm和G/Dsah均能正常生长(图 2A),说明敲除spe2、msm1和sah1不会导致工程菌变成营养缺陷型。

|
| 图 2 毕赤酵母工程菌在MD (A)、BMGY (B)、BMMY (C)和BSM (D)培养基中的生长 Figure 2 Cell growth of engineered strains on MD medium (A) and in BMGY medium (B), BMMY medium (C), BSM medium (D). |
|
|
以甘油为碳源时检测了工程菌在BMGY培养基中的生长情况。从MD平板上挑取单菌落接种至3 mLYPG液体培养基中培养过夜,再取1 mL培养液置于25 mL BMGY中进行培养。结果显示,与出发菌GS115/DS16相比,工程菌G/Dspe、G/Dmsm和G/Dsah的生长无显著差异;培养16 h菌体生长进入平稳期(图 2B)。
在甲醇诱导培养基中(BMMY和BSM)检测了工程菌的生长。取BMGY培养16 h的培养液,于4 000 r/min离心5 min,弃上清,菌体重悬于25 mL BMMY或BSM中。工程菌于30 ℃、220 r/min和甲醇诱导条件下培养96 h,每隔12 h添加甲醇至终浓度为1.2% (体积分数)。在甲醇诱导培养基BMMY中,与出发菌株GS115/DS16相比,工程菌G/Dsah生长较缓慢,G/Dspe和G/Dmsm无显著差异(图 2C);在基础盐培养基BSM中,工程菌与出发菌的生长趋势相似且未出现停滞期,菌体生长呈持续上升的趋势,菌体量显著高于BMMY培养条件下(图 2D)。因此,后继发酵SAM培养基采用生长较好的BSM培养基。
2.3 阻断SAM分解代谢途径对其产量的影响从工程菌分别挑选6个阳性转化子在摇瓶中检测其SAM产量,在基础盐培养基BSM中发酵,甲醇诱导12 h后,每隔24 h添加L-Met至终浓度为0.10% (质量体积分数),诱导96 h后测定各工程菌的菌体量和胞内SAM浓度。
与对照菌GS115/DS16的单位菌体干重SAM产量(98.8 mg/g-DCW)相比,工程菌G/Dspe、G/Dsah和G/Dmsm的SAM产量分别提高了29.3%、55.6%和24.8%,达到127.7、153.7和123.3 mg/g-DCW;其中G/Dsah的SAM产量提高幅度最大,提高了55.6% (图 3)。检测不同工程菌胞内的MAT酶活,与对照菌GS115/DS16的MAT酶活[(236.5±19.4) U/g-DCW]相比,各工程菌的MAT酶活[G/Dspe:(244.7±20.8) U/g-DCW;G/Dsah:(228.4±18.6) U/g DCW;G/Dmsm:(240.2±18.5) U/g-DCW]无显著性差异,说明工程菌SAM产量的提高并非其MAT酶活提高造成。

|
| 图 3 工程菌G/Dspe、G/Dsah和G/Dmsm转化子的SAM产量盒须图 Figure 3 Box-plot representation of SAM specific production of G/Dspe, G/Dsah and G/Dmsm transformants. 每个工程菌挑选6个转化子进行摇瓶发酵,诱导96 h,将转化子的单位菌体干重SAM产量做成盒须图. 在盒须图中,—:样本中最大值和最小值;□:平均值;长方形盒子内的样本数占75%. 分别对工程菌和对照菌GS115/DS16的数据进行t检验显著性分析,*:P < 0.05 Six transformants from each engineered stain were cultivated in shake flask for 96 h. Box-plot is used to show the distribution of SAM specific production. In box plot, —: The maximum and minimum values in the sample; □: Average value; the box represents 75% of the sample size. *: P < 0.05 indicate significant differences between engineered strains and parental strain GS115/DS16 by Student's t-test. |
|
|
L-Met直接参与细胞中SAM的合成,利用微生物合成SAM时,优化L-Met添加策略可提高SAM产量。但额外添加的L-Met浓度过高时会抑制MAT酶活,对菌体生长也有影响[20]。因此,我们研究了不同的L-Met添加量(0.06%和0.10%)对工程菌SAM产量和菌体生长的影响。
2.4.1 L-Met添加量对工程菌G/Dspe合成SAM的影响在基础盐培养基BSM中,甲醇诱导96 h,诱导期间每隔24 h加入L-Met至终浓度为0.06%或0.10% (质量体积分数)。不同的L-Met添加量对工程菌G/Dspe的生长和单位菌体干重SAM产量无显著影响(图 4)。

|
| 图 4 L-Met添加量对工程菌G/Dspe生长(A)和SAM产量(B)的影响 Figure 4 The influence of L-Met addition on cell growth (A) and SAM accumulation (B) of engineered strain G/Dspe. |
|
|
不同L-Met添加量条件下,工程菌G/Dsah的生长无显著差异。当L-Met添加量为0.10%时,G/Dsah的单位菌体干重SAM产量为(155.5±9.5) mg/g-DCW。L-Met添加量为0.06%时,G/Dsah的单位菌体干重SAM产量达到(196.6±12.2) mg/g-DCW,比0.10% L-Met添加量时提高了26.4% (图 5)。因此,0.06%的L-Met添加量更适合G/Dsah生产SAM。

|
| 图 5 L-Met添加量对工程菌G/Dsah的生长(A)和SAM产量(B)的影响 Figure 5 The influence of L-Met addition on cell growth (A) and SAM accumulation (B) of engineered strain G/Dsah. 对数据进行t检验显著性分析. **:P < 0.01 **: P < 0.01 indicate significant differences between the results on different L-Met concentration by Student's t-test. |
|
|
当L-Met添加量为0.06%时,在发酵后期(72−96 h),工程菌G/Dmsm的生物量和单位菌体干重SAM产量显著高于L-Met添加量为0.10%时(图 6)。当L-Met添加量为0.10%时,G/Dmsm的单位菌体干重SAM产量为(136.8±7.1) mg/g-DCW,L-Met添加量为0.06%时,G/Dmsm的菌体量和单位菌体干重SAM产量[(176.4±9.2) mg/g-DCW]分别比0.10% L-Met添加量时提高了17.2%和28.9%。检测了工程菌G/Dmsm胞内的MAT酶活,当L-Met添加量为0.06%时MAT酶活为(274.4±21.3) U/g-DCW,较优化前[(240.2±18.5) U/g-DCW]提高了14.2%。结果表明,0.06% L-Met添加量比0.10% L-Met添加量更适合G/Dmsm生产SAM。

|
| 图 6 L-Met添加量对工程菌G/Dmsm生长(A)和SAM产量(B)的影响 Figure 6 The influence of L-Met addition on cell growth (A) and SAM accumulation (B) of engineered strain G/Dmsm. 对数据进行t检验显著性分析. *:P < 0.05;**:P < 0.01 *: P < 0.05; **: P < 0.01 indicate significant differences between the results on different L-Met concentration by Student's t-test. |
|
|
本研究通过敲除重组菌GS115/DS16 (过表达高活性MAT的毕赤酵母重组菌)的S-腺苷同型半胱氨酸水解酶基因sah1、S-腺苷甲硫氨酸脱羧酶基因spe2和L-甲硫氨酰tRNA合酶基因msm1,分别抑制了SAM经转甲基向AdoHcy转化、经脱羧向脱羧S-腺苷甲硫氨酸转化以及向L-甲硫氨酰-tRNA转化,减少SAM在胞内的消耗利用,提高了工程菌积累SAM的能力。通过优化L-Met添加量,进一步提高了工程菌G/Dmsm和G/Dsah的SAM产量。
在毕赤酵母中,敲除spe2、sah1和msm1不会造成菌株的营养缺陷(图 2A)。SAM脱羧转化为脱羧S-腺苷甲硫氨酸是多胺合成的重要途径[21]。在粟酒裂殖酵母(Schizosaccharomyces pombe)和酿酒酵母(Saccharomyces cerevisiae)中,敲除S-腺苷甲硫氨酸脱羧酶基因spe2后,细胞不能合成亚精胺或精胺,耗尽胞内多胺后停止成长,会造成亚精胺营养缺陷型,培养基中需额外添加亚精胺菌体才能生长[22-23]。但我们的结果显示,敲除spe2后,工程菌G/Dspe在BMMY培养基或BSM基础盐培养基中都能正常生长。与对照菌GS115/DS16相比,G/Dspe菌株的单位菌体干重SAM产量提高了29.3%。
S-腺苷高半胱氨酸(AdoHcy)作为SAM众多转甲基化反应的共同产物,胞内的SAM/AdoHcy值处于一个稳定水平。SAH的功能很可能与L-Met/半胱氨酸的体内平衡有关,但敲除sah1并不影响毕赤酵母的正常生长;只有在缺乏硫同化的情况下,sah1在酵母中才是必需的[24]。合成培养基BMMY中,与出发菌株GS115/DS16相比,G/Dsah菌株的生长较缓慢;而在基础盐培养基BSM中,G/Dsah菌株的生长与对照菌无显著差异(图 2C、2D)。这很可能是因为与合成培养基BMMY相比,基础盐培养基BSM中含有更丰富的硫,可以补偿工程菌由于sah1敲除后不能进行的硫循环。sah1敲除后抑制了G/Dsah胞内的转甲基和转巯基作用,一定程度上减少了SAM的转化利用。因此,G/Dsah菌株的单位菌体干重SAM产量提高幅度最大,提高了55.6%。类似的研究发现,在日本清酒酵母中引入突变的sah1并敲除野生型SAH1后SAM产量提高了2.5倍[25]。
参与SAM合成的底物L-Met经L-甲硫氨酰tRNA合酶MSM1催化[10],先形成L-甲硫氨酰-tRNA,再生成N-甲酰-甲硫氨酰-tRNA,参与生理反应。敲除msm1阻断了工程菌G/Dmsm由L-Met生成L-甲硫氨酰-tRNA的途径,一定程度上提高了胞内的L-Met浓度,但并未影响其正常生长。在BMMY和BSM培养基中,G/Dmsm菌株和对照菌的生长曲线基本持平。细胞内L-Met浓度增加有利于SAM的合成,与对照菌DS16相比,G/Dmsm菌株的单位菌体干重SAM产量提高了24.8%。
L-Met是细胞内重要的含硫化合物,参与许多重要的生理代谢,包括合成蛋白质和活性甲基供体SAM。在植物的相关研究中发现,Lemna pausicostata将细胞中80%的L-Met用于合成SAM[26]。利用微生物合成SAM时,优化发酵培养中的L-Met添加量可提高SAM产量并降低发酵成本。但添加的L-Met浓度过高会抑制MAT酶活和菌体生长[20]。与出发菌株GS115/DS16相比,工程菌G/Dspe、G/Dmsm和G/Dsah胞内的L-Met浓度很可能会有不同程度的增加。发酵过程中添加相同量的L-Met,各工程菌的L-Met消耗量可能会由于胞内原有积累程度的不同而改变。敲除sah1后可能会影响到SAM合成反应底物L-Met的循环,从而影响SAM的积累。L-Met添加量为0.06%时,G/Dsah的单位菌体干重SAM产量比0.10% L-Met添加量时提高了26.4%。MSM1直接和L-Met代谢相关,敲除msm1很可能会提高胞内的L-Met浓度。L-Met添加量为0.06%时,G/Dmsm的菌体量、单位菌体干重SAM产量和MAT酶活水平比0.10% L-Met添加量时分别提高了17.2%、28.9%和14.2%。这说明低浓度的L-Met添加不仅有利于G/Dmsm菌体的生长,而且对MAT酶活和SAM产量也有促进作用。L-Met作为底物,是SAM生物合成的瓶颈之一,发酵过程要额外补加;当L-Met添加量较高时(0.15%),酵母的TCA循环和氧化磷酸化作用被显著抑制,胞内ATP也基本被耗尽[13]。ATP的减少不利于细胞的增殖,也很可能成为SAM生产的限制步骤。因此,相较于高浓度的L-Met添加量,在保证底物供应的情况下,低浓度L-Met添加量(0.06%)更适于工程菌SAM的生产。
本研究通过阻断毕赤酵母胞内SAM的转化利用途径,进一步提高了工程菌积累SAM的能力。与出发菌株相比,工程菌G/Dspe、G/Dsah和G/Dmsm的单位菌体干重SAM产量分别提高了29.3%、55.6%和24.8%。通过优化L-Met添加量,G/Dsah和G/Dmsm的单位菌体干重SAM产量分别达到(196.6±12.2) mg/g-DCW和(176.4±9.2) mg/g-DCW,是出发菌的2.0倍和1.8倍。通过代谢工程改造对SAM代谢途径相关基因进行多基因的过表达或敲除,毕赤酵母和酿酒酵母的单位菌体干重SAM产量最高可达到186.3 mg/g-DCW[4]和455.1 mg/g-DCW[3]。本研究构建的工程菌G/Dsah,在优化L-Met添加量后,单位菌体干重SAM产量达到目前报道的毕赤酵母最高水平;单基因敲除工程菌的SAM产量较出发菌的均有提高,双基因或三基因组合敲除后,工程菌的SAM产量很可能会进一步提升。由于各工程菌的基因型不同,其生理生化特性存在差异,SAM发酵的最优条件也不尽相同,因此,经发酵调控优化后工程菌的SAM产量还有进一步提升的潜力。
REFERENCES
| [1] |
MISCHOULON D, ALPERT JE, ARNING E, BOTTIGLIERI T, FAVA M, PAPAKOSTAS GI. Bioavailability of S-adenosyl methionine and impact on response in a randomized, double-blind, placebo-controlled trial in major depressive disorder[J]. The Journal of Clinical Psychiatry, 2012, 73(6): 843-848. DOI:10.4088/JCP.11m07139 |
| [2] |
PARASHAR S, CHEISHVILI D, ARAKELIAN A, HUSSAIN Z, TANVIR I, KHAN HA, SZYF M, RABBANI SA. S-adenosylmethionine blocks osteosarcoma cells proliferation and invasion in vitro and tumor metastasis in vivo: therapeutic and diagnostic clinical applications[J]. Cancer Medicine, 2015, 4(5): 732-744. DOI:10.1002/cam4.386 |
| [3] |
CHEN HL, ZHU NQ, WANG Y, GAO XX, SONG YH, ZHENG J, PENG JP, ZHANG X. Increasing glycolysis by deletion of kcs1 and arg82 improved S-adenosyl-L-methionine production in Saccharomyces cerevisiae[J]. AMB Express, 2021, 11(1): 20. DOI:10.1186/s13568-021-01179-8 |
| [4] |
QIN XL, LU JJ, ZHANG Y, WU XL, QIAO XF, WANG ZP, CHU J, QIAN JC. Engineering Pichia pastoris to improve S-adenosyl-L-methionine production using systems metabolic strategies[J]. Biotechnology and Bioengineering, 2020, 117(5): 1436-1445. DOI:10.1002/bit.27300 |
| [5] |
XU CT, SHI ZW, SHAO JQ, YU CK, XU ZN. Metabolic engineering of Lactococcus lactis for high level accumulation of glutathione and S-adenosyl-L-methionine[J]. World Journal of Microbiology & Biotechnology, 2019, 35(12): 185. |
| [6] |
HAYAKAWA K, MATSUDA F, SHIMIZU H. 13 C-metabolic flux analysis of ethanol-assimilating Saccharomyces cerevisiae for S-adenosyl-L-methionine production[J]. Microbial Cell Factories, 2018, 17(1): 82. DOI:10.1186/s12934-018-0935-6 |
| [7] |
GODSPOWER H, QIAO ZN, XU MJ, RAO ZM. Expression and catalytic application of S-adenosylmethionine synthase from different sources in Escherichia coli[J]. Food and Fermentation Industries, 2022. Hero Nmeri Godspower, 乔郅钠, 徐美娟, 饶志明. 不同来源S-腺苷甲硫氨酸合酶在大肠杆菌中的表达及催化应用[J]. 食品与发酵工业, 2022. DOI:10.13995/j.cnki.11-1802/ts.031256 |
| [8] |
QIN XL, QIAN JC, YAO GF, ZHUANG YP, ZHANG SL, CHU J. GAP promoter library for fine-tuning of gene expression in Pichia pastoris[J]. Applied and Environmental Microbiology, 2011, 77(11): 3600-3608. DOI:10.1128/AEM.02843-10 |
| [9] |
CHEN HL, WANG ZL, CAI HB, ZHOU CL. Progress in the microbial production of S-adenosyl-L-methionine[J]. World Journal of Microbiology & Biotechnology, 2016, 32(9): 153. |
| [10] |
SERRE L, VERDON G, CHOINOWSKI T, HERVOUET N, RISLER JL, ZELWER C. How methionyl-tRNA synthetase creates its amino acid recognition pocket upon L-methionine binding[J]. Journal of Molecular Biology, 2001, 306(4): 863-876. DOI:10.1006/jmbi.2001.4408 |
| [11] |
JIANG LL, WU L, XU HX, HUANG JL, ZHANG YJ, XU Q, YANG Y. Construction of ATP synthetic strain and its application in the production of S-adenosylmethionine[J]. Biotechnology Bulletin, 2019, 35(6): 221-226. (in Chinese) 江林林, 吴磊, 许海霞, 黄坚丽, 张永进, 徐期, 杨勇. ATP合成菌株的构建及用于联合生产S-腺苷甲硫氨酸[J]. 生物技术通报, 2019, 35(6): 221-226. DOI:10.13560/j.cnki.biotech.bull.1985.2018-0977 |
| [12] |
ZHENG JR, CHEN LF, YANG SY, ZHU JR. Synthesis of S-adenosylmethionine from adenine in constructing engineered strain and the screening of high yield strain[J]. Food and Fermentation Industries, 2014, 40(9): 23-28. (in Chinese) 郑计瑞, 陈丽芬, 杨善岩, 朱家荣. 腺嘌呤合成腺苷甲硫氨酸的工程菌构建及其高产株筛选[J]. 食品与发酵工业, 2014, 40(9): 23-28. |
| [13] |
KENSHI H, FUMIO M, HIROSHI S. Metabolome analysis of Saccharomyces cerevisiae and optimization of culture medium for S-adenosyl-L-methionine production[J]. AMB Express, 2016, 6(1): 38. DOI:10.1186/s13568-016-0210-3 |
| [14] |
HU H, QIAN JC, CHU J, WANG YH, ZHUANG YP, ZHANG SL. Optimization of L: -methionine feeding strategy for improving S-adenosyl-L: -methionine production by methionine adenosyltransferase overexpressed Pichia pastoris[J]. Applied Microbiology and Biotechnology, 2009, 83(6): 1105-1114. DOI:10.1007/s00253-009-1975-y |
| [15] |
LIU W, TANG DD, SHI R, LIAN JZ, HUANG L, CAI J, XU ZN. Efficient production of S-adenosyl-L-methionine from dl-methionine in metabolic engineered Saccharomyces cerevisiae[J]. Biotechnology and Bioengineering, 2019, 116(12): 3312-3323. DOI:10.1002/bit.27157 |
| [16] |
RAVI KANT H, BALAMURALI M, MEENAKSHISUNDARAM S. Enhancing precursors availability in Pichia pastoris for the overproduction of S-adenosyl-L-methionine employing molecular strategies with process tuning[J]. Journal of Biotechnology, 2014, 188: 112-121. DOI:10.1016/j.jbiotec.2014.08.017 |
| [17] |
CHEN H, WANG Z, WANG Z, DOU J, ZHOU C. Improving methionine and ATP availability by MET6 and SAM2 co-expression combined with sodium citrate feeding enhanced SAM accumulation in Saccharomyces cerevisiae[J]. World Journal of Microbiology & Biotechnology, 2016, 32(4): 56. |
| [18] |
HU H, QIAN JC, CHU J, WANG Y, ZHUANG YP, ZHANG SL. DNA shuffling of methionine adenosyltransferase gene leads to improved S-adenosyl-L-methionine production in Pichia pastoris[J]. Journal of Biotechnology, 2009, 141(3/4): 97-103. |
| [19] |
YAO GF, QIN XL, CHU J, WU XL, QIAN JC. Expression, purification, and characterization of a recombinant methionine adenosyltransferase pDS16 in Pichia pastoris[J]. Applied Biochemistry and Biotechnology, 2014, 172(3): 1241-1253. DOI:10.1007/s12010-013-0594-3 |
| [20] |
HE JY, DENG JJ, ZHENG YH, GU J. A synergistic effect on the production of S-adenosyl-L-methionine in Pichia pastoris by knocking in of S-adenosyl-L-methionine synthase and knocking out of cystathionine-beta synthase[J]. Journal of Biotechnology, 2006, 126(4): 519-527. DOI:10.1016/j.jbiotec.2006.05.009 |
| [21] |
BALASUNDARAM D, DINMAN JD, TABOR CW, TABOR H. SPE1 and SPE2: two essential genes in the biosynthesis of polyamines that modulate +1 ribosomal frameshifting in Saccharomyces cerevisiae[J]. International Journal of Molecular Sciences, 1994, 176(22): 7126-7128. |
| [22] |
CHATTOPADHYAY MANAS K, WHITE TC, HERBERT T. Absolute requirement of spermidine for growth and cell cycle progression of fission yeast (Schizosaccharomyces pombe)[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(16): 10330-10334. |
| [23] |
CHATTOPADHYAY MK, TABOR CW, TABOR H. Spermidine but not spermine is essential for hypusine biosynthesis and growth in Saccharomyces cerevisiae: spermine is converted to spermidine in vivo by the FMS1-amine oxidase[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(24): 13869-13874. |
| [24] |
NERMINA M, INGO S, HEIMO W, GERALD R, KOHLWEIN SEPP D, OKSANA T. S-adenosyl-L-homocysteine hydrolase, key enzyme of methylation metabolism, regulates phosphatidylcholine synthesis and triacylglycerol homeostasis in yeast: implications for homocysteine as a risk factor of atherosclerosis[J]. The Journal of Biological Chemistry, 2008, 283(35): 23989-23999. |
| [25] |
ANO A, SUEHIRO D, CHA-AIM K, ARITOMI K, PHONIMDAENG P, NONTASO N, HOSHIDA H, MIZUNUMA M, MIYAKAWA T, AKADA R. Combinatorial gene overexpression and recessive mutant gene introduction in sake yeast[J]. Bioscience, Biotechnology, and Biochemistry, 2009, 73(3): 633-640. |
| [26] |
STÉPHANE R, BLOCK MARYSE A, PASCAL R, SAMUEL J, GILLES C, FABRICE R, ROLAND D. Methionine metabolism in plants: chloroplasts are autonomous for de novo methionine synthesis and can import S-adenosylmethionine from the cytosol[J]. The Journal of Biological Chemistry, 2004, 279(21): 22548-22557. |